結果及び考察303-2
|
|
|
- さみら たかひ
- 7 years ago
- Views:
Transcription
1 3. ルーメンフローラ解析 1) 生菌剤 (BLCS) の影響 1 目的 BLCS ルナシータは 日本エメラル株式会社により 米ぬか 小麦フスマ 大豆粕などを吸着剤として枯草菌 乳酸菌 酵母などの微生物を混合発酵させて製造されたものである これを牛および羊に投与することにより ルーメンでのメタン生成が抑制できることが報告されている ( 高浦ら 第 106 回日本畜産学会 ) しかし BLCS ルナシータの効果がメタン生成菌に対して直接的に作用しているのか あるいは他のルーメン細菌を介した間接的な作用であるかは明らかにされていない そこで本試験では BLCS ルナシータを羊に投与することにより生じるルーメンフローラの変化 ( メタン生成菌数の低下 BLCS ルナシータに含まれる細菌の増加 水素消費細菌の増加など ) を明らかにすることを目的とした 2 試験対象サンプル試験対象サンプルは帯広畜産大学殿から提供を受けたルーメン液と BLCS ルナシータである ルーメン液は給与飼料のみを投与 ( 対照区 ) モネンシンを 1 日当たり 20mg 投与 ( モネンシン区 ) BLCS ルナシータを 1 日当たり 4g 投与 (4g 区 ) および BLCS ルナシータを 1 日当たり 8g 投与 (8g 区 ) したチェビオット種去勢羊から採取されたものである 羊への給与条件は以下の通りである 対照区モネンシン区 4g 区 BLCS 8g 区 BLCS 給与飼料のみを投与モネンシンを 1 日当たり 20mg 投与ルナシータを 1 日当たり 4g 投与ルナシータを 1 日当たり 8g 投与 給与期間を 20 日とし 14 日間の間隔を空けたものを 1 サイクルとして Ⅰ 期から Ⅳ 期ま での 4 サイクルの試験にわたり ルーメン液の採取が行なわれた 3 試験方法本試験では DGGE 法 T-RFLP 法および MPN-PCR 法の三種類を用いた DGGE( 変性グラジエントゲル電気泳動 ) 法を用いるルーメンフローラの解析対象として 真正細菌 古細菌 真核生物とした 真正細菌の解析により BLCS ルナシータ投与による BLCS ルナシータに含まれる細菌の出現および水素消費細菌の増減を把握することを目的とした メタン生成菌を含む古細菌の解析により ルーメン内に含まれるメタン生成菌の種類を特定することを目的とした 酵母を含む真核生物の解析により BLCS ルナシータに含まれる酵母がルーメン内における増加について明らかにすることを目的とした T-RFLP( 制限酵素末端断片長解析 ) 法を用いるルーメンフローラの解析対象は真正細菌とし
2 た DGGE 法の解析結果について 真正細菌の増減を定量的に裏付けることを目的として 実施した MPN-PCR 法 ( 最確数 -PCR) により BLCS ルナシータがメタン生成菌数の低減に効果を有 しているかを定量的に確認することを目的とした 4 試験 1.PCR-DGGE 法によるルーメンフローラの解析 4-1 PCR-DGGE 法のフロー 図 4-1 に PCR-DGGE 法による微生物群集構造解析のフローを示す 1. ルーメン液からの DNA 抽出 精製 2. PCR 反応 ( プライマー :GC クランプ付 ) 1)a,b,c 真正細菌 古細菌 真核生物を対象とした PCR 条件 (1 回目 ) 2)a,b,c 真正細菌 古細菌 真核生物を対象とした PCR 反応 (GC クランプ付き ) 3. アガロース電気泳動による 16S rdna の増幅の確認 DNA バンドが単一になる まで数回繰り返す 4. DGGE 解析 5. DNA バンド切り出しと DNA 抽出 精製 6. PCR 反応 ( プライマー :GC クランプの除去 ) 7. PCR 産物の精製 8. シーケンス解析 10. ホモロジー解析 ( 微生物種の同定 ) 図 4-1 PCR-DGGE 法による微生物群集構造解析のフロー 4-2 ルーメン液からの DNA 抽出 精製ルーメン液には DNA 抽出が困難な細菌および PCR 阻害物質が存在すると考えられる このため 本試験では強固な細胞壁を備えた微生物の物理的破壊工程 界面活性剤およびその他薬剤を 65 で作用させる DNA 抽出工程 腐植物質 ( 或いは前駆物質であるリグニン ) の除去工程を含む方法 (Yoshida et al. 2005) を用いた
3 4-3 PCR 反応解析対象は真正細菌 古細菌および真核生物とした 表 4-1 に真正細菌 古細菌および真核生物を対象としたプライマーの配列を示した 表 4-1 真正細菌を対象とした PCR に用いたプライマーの配列
4 4-3-1a) 真正細菌を対象とした PCR 条件 (1 回目 ) 抽出した DNA を鋳型として 真正細菌用のプライマー (341F 907R) を用いて 16SrDNA を増幅した PCR 条件を図 4-2 に示した [ PCR 反応 (Primer : 341F, 907R) ] 96 5min 96 1min 55 30sec 25cycle 72 1min 72 5min 4 図 4-2 真正細菌の 16SDNA を標的とした PCR 反応条件 4-3-2a) 真正細菌を対象とした PCR 反応 (GC クランプ付き ) DGGE ゲルを用いた電気泳動を行うため GC クランプ付 (GC に富む配列 ) プライマーを用いた PCR を行った 図 4-2 で得られた PCR 産物を鋳型として 真正細菌用 GC クランプ付きプライマー (341FGC,534R) を用いて PCR を行い ( 図 4-3) アガロースゲル電気泳動にて PCR 産物の増幅を確認した [ PCR 反応 (Primer : 341FGC, 534R) ] 96 5min 96 1min 55 30sec 25cycle 72 30sec 72 5min 4 図 4-3 真正細菌の 16SDNA を標的とした PCR 反応条件 (GC クランプ付 )
5 4-3-1b) 古細菌を対象とした PCR 条件 ( オプション ) 抽出した DNA を鋳型として 古細菌用のプライマー (46F 1100R) を用いて 16S rdna を 増幅した PCR 条件を図 4-4 に示した PCR 反応 (Primer: 46F, 1100R) 94 10min 92 1min sec 30cycle 72 1min 72 6min 4 図 4-4 古細菌の 16S rdna を標的とした PCR 反応条件 4-3-2b) 古細菌を対象とした PCR 反応 (GC クランプ付き ) ( オプション ) DGGE ゲルを用いた電気泳動を行うため 真正細菌と同様 GC クランプ付プライマーを用いた PCR を行った 図 4-4 で得られた PCR 産物を鋳型として 古細菌用 GC クランプ付きプライマー (340FGC 519R) を用いて PCR を行い ( 図 4-5) アガロースゲル電気泳動にて PCR 産物の増幅を確認した PCR 反応 (Primer: 340F, 519R) 94 10min 92 1min 65 30sec 30cycle 72 1min 72 6min 4 図 4-5 古細菌の 16SDNA を標的とした PCR 反応条件 (GC クランプ付 )
6 4-3-1c) 真核生物を対象とした PCR 条件 ( オプション ) 抽出した DNA を鋳型として 真核生物用のプライマー (NS1 ITS4) を用いて 18S rdna を増幅した PCR 条件を図 4-6 に示した PCR 反応 (Primer: NS1, ITS4) 96 5min 96 1min 40 1min 30cycle 72 1min 72 5min 4 図 4-6 真核生物の 18S rdna を標的とした PCR 反応条件 4-3-2c) 真核生物を対象とした PCR 反応 (GC クランプ付き ) ( オプション ) DGGE ゲルを用いた電気泳動を行うため 真正細菌 古細菌と同様 GC クランプ付プライマーを用いた PCR を行った 図 4-6 で得られた PCR 産物を鋳型として 真核生物用 GC クランプ付きプライマー (NS7-GC F1Ra) を用いて PCR を行い ( 図 4-7) アガロースゲル電気泳動にて PCR 産物の増幅を確認した PCR 反応 (Primer: NS7-GC, F1Ra) 96 5min 96 1min 60 30sec 30cycle 72 30sec 72 5min 4 図 4-7 真核生物類をターゲットとした nested PCR 条件
7 4-4 DGGE 解析広範囲のバンドが得られるように ゲル中の DNA 変性剤の濃度勾配を 35-60%( 真正細菌 古細菌 ) および 30-70%( 真核生物類 )( 変性剤 100% とは 7M 尿素 40% ホルムアミドに相当 ) として DGGE 解析を行った また 重合剤の濃度は 低濃度に設定し ゲルを 4 時間固化させることにより グラジエントの精度を高めたゲルを作成した さらに バンドの分離能を高めるため 電圧を低くし 泳動時間を長く設定した ( 表 4-2) 表 4-2 DGGE ゲル濃度と泳動条件 ポリアクリルアミドゲル濃度 :10%( 真正細菌 古細菌 ) :6%( 真核生物 ) 変性剤濃度勾配 :35-60%( 真正細菌 古細菌 ) :30-70%( 真核生物 ) 重合剤濃度 10% Ammonium Persulfate :0.7% TEMED :0.07% 泳動電圧 :70V 泳動時間 :960min 泳動温度 : DNA バンドの切り出しと DNA 抽出 精製電気泳動後 エチジウムブロマイドで染色を行い 脱染色後 UV トランスイルミネーター上でバンドを検出した 検出したバンドを滅菌されたメスで切り出し 1.5ml チューブに入れた 切り出したゲルに 100μl の TE buffer を加え 35 で一晩振とうさせた後 エタノール沈殿法により DNA の抽出 精製を行った DGGE ゲルからのバンド切り出し DNA シーケンスを行う基準を以下に示す BLCS ルナシータ投与区において増減の見られるバンド BLCS ルナシータに含まれるバンドに近いもの とした なお 分離能が低く 複数のバンドが混在している可能性のあるバンドについては バンドが単一になるまで GC クランプ付 PCR および DGGE 解析を繰り返した
8 4-6 PCR 反応 (GC クランプの除去 ) 各バンドから抽出した DNA を鋳型として シーケンス反応を行うため GC クランプを 外す PCR を行った 1) 真正細菌の GC クランプ無し PCR 真正細菌の GC クランプを外す PCR の条件を図 4-8 に示した PCR 反応 (Primer : 341F, 534R) 96 5min 96 1min 55 30sec 25cycle 72 30sec 72 5min 4 図 4-8 GC クランプの除去に用いた PCR 反応条件 ( 真正細菌 ) 2) 古細菌の GC クランプ無し PCR( オプション ) 古細菌の GC クランプを外す PCR の条件を図 4-9 に示した PCR 反応 (Primer : 340F, 519R) 94 5min 92 1min 65 30sec 30cycle 72 30sec 72 5min 4 図 4-9 GC クランプの除去に用いた PCR 反応条件 ( 古細菌 )
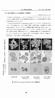
9 3) 真核生物の GC クランプ無し PCR( オプション ) 真核生物の GC クランプを外す PCR の条件を図 4-10 に示した PCR 反応 (Primer : NS7F,F1Ra) 96 5min 96 30sec 50 30sec 30cycle 72 30sec 72 5min 4 図 4-10 GC クランプの除去に用いた PCR 反応条件 ( 真核生物 ) 4-7 PCR 産物の精製アガロースゲル電気泳動により PCR 産物を確認し 次いで アガロースから目的バンドを切り出して 精製キット (QIAquickⅡ Purification Kit; QIAGEN 社製 ) により PCR 産物を精製した 4-8 シーケンス解析 4-7 で精製を行った PCR 産物を鋳型として ダイターミネーター法 (Big Dye Terminator v1.1 Cycle Sequencing Kit;ABI 社製 ) によりシーケンシング反応を行った シーケンス解析は ABI Prism310 Genetic Analyzer(ABI 社製 ) を用いて行った 4-9 ホモロジー解析 ( 微生物種の同定 ) 解読した各バンドの DNA シーケンスを基に インターネットを利用した FASTA 検索により DDBJ ( 日本 ) EMBL( ヨーロッパ ) のデータベースから微生物種の同定を行った 4-10 PCR-DGGE 法による解析結果 4-11 真正細菌の DGGE 解析結果真正細菌の DGGE バンドパターンを図 4-11(Ⅰ 期およびⅡ 期 ) および 4-12(Ⅲ 期およびⅣ 期 ) に示した また 塩基配列を決定したバンドの位置を四角で囲った

10 図 4-11 真正細菌 DGGE 解析結果 (Ⅰ 期 Ⅱ 期 ) ( 白黒反転画像 ) BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示 す

11 図 4-12 真正細菌 DGGE 解析結果 (Ⅲ 期 Ⅳ 期 ) ( 白黒反転画像 ) BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示 す
12 4-12 古細菌の DGGE 解析結果 ( オプション ) 古細菌の DGGE バンドパターンを図 4-13 に示した 蛍光輝度のやや強いバンドが一本と極めて薄いバンドが 2 本見られた なお 塩基配列を決定したバンドの位置を四角で囲った 図 4-13 古細菌 DGGE 解析結果 ( 白黒反転画像 ) BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示 す
 BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示")
13 4-13 真核生物の DGGE 解析結果 ( オプション ) 真核生物の DGGE 染色画像を図 4-14 に示した バンドが認められなかったため 塩基配 列の同定は行わなかった 図 4-14 真核生物 DGGE 解析結果 ( 白黒反転画像 ) BLCS: BLCS ルナシータ, con: 対照区, mone: モネンシン区, 4g: 4g 区, 8g: 8g 区を示 す
14 4-14 シーケンス解析結果シーケンス解析の結果決定した真正細菌の塩基配列を表 4-3 に 古細菌の塩基配列を表 4-4 に示した 表 4-3 決定した塩基配列 ( 真正細菌 )

15 表 4-4 決定した塩基配列 ( 古細菌 )
16 4-15. ホモロジー解析結果 ( 微生物種の同定 ) 各バンドの塩基配列に基づいて DDBJ-FASTA 検索を行い ホモロジーの高かった上位 5 種を表に示した ( 真正細菌 : 表 4-5 古細菌 : 表 4-6) また EMBL-FASTA 検索も行い ホモロジーの高い上位 5 種を表 4-7( 真正細菌 ) および表 4-8( 古細菌 ) に示した また それらの Identity( 相同性 ;% で表す ) と Overlap した領域 (nt: 塩基対数 ) を表記した DDBJ-FASTA と EMBL-FASTA の結果を比較すると 全ての結果において EMBL-FASTA の方が相同性が高いあるいは同等であった 表 4-5 真正細菌の同定結果 (DDBJ-FASTA)
17
18
19
20
21 表 4-6 古細菌の同定結果 (DDBJ-FASTA)
22 表 4-7 真正細菌の同定結果 (EMBL-FASTA)
23
24
25

26 表 4-8 古細菌の同定結果 (EMBL-FASTA) 4-16 同定した細菌の生態的特徴表 4-9 に同定した真正細菌の生態的特徴を示し 表 4-10 に同定した古細菌の生態的特徴を示した なお 4-15 に示した通り DDBJ-FASTA と比較して高い相同性の結果が得られた EMBL-FASTA による結果を用いた 表 4-9 真正細菌の生態特徴 (EMBL-FASTA の結果に基づく )

27

28

29

30

31

32
33 表 4-10 古細菌の生態特徴 (EMBL-FASTA の結果に基づく )
34 4-17 真正細菌の出現結果表 4-11 に DGGE 解析により得られたバンド輝度から試料に含まれる微生物量を判定した結果を示した 細菌名の欄には EMBL-FASTA のデータベース上に存在する細菌の 16S rdna の配列データと相同性が最も高いものを示した

35 表 4-11 真正細菌の出現状況 BL: BLCS ルナシータ, con: 対照区, mo: モネンシン区, 4g 区 8g 区 : 多い, : 少ない, : 極めて少ないまたは無い
36 DGGE 法による解析の結果 BLCS ルナシータに含まれる細菌は BLCS ルナシータを投与した羊のルーメン液中からは検出されなかった 第 Ⅰ 期においては 対照区と比較して BLCS ルナシータ 4g 区で Rumen bacterium および Clostridium thermoscinogenes のバンドの蛍光輝度が向上し 8g 区において Rumen bacterium Clostridium thermoscinogenes Clostridium leptum および Succiniclasticum ruminis のバンドの蛍光輝度の向上が認められた 一方 モネンシン区においては対照区と比較して大きな変化は見られなかった 第 Ⅱ 期においては 対照区と比較して 4g 区で Rumen bacterium および Clostridium thermoscinogenes のバンドの蛍光輝度の向上が見られ 8g 区では Rumen bacterium Clostridium thermoscinoges および Succiniclasticum ruminis のバンドの蛍光輝度の向上が認められた モネンシン区においては対照区と比較して Clostridium thermoscinogenes Clostridium leptum Swine manure bacterium のバンドの蛍光輝度の向上が認められた 第 Ⅲ 期においては 対照区と比較して 4g 区で Clostridium thermoscinogenes のバンドの蛍光輝度の向上が見られ Clostridium leptum のバンドの蛍光輝度の低下が認められた BLCS ルナシータ 8g 区では Clostridium thermoscinogenes のバンドの蛍光輝度の向上が見られ Rumen bacterium Clostridium leptum および Quinella ovalis のバンドの蛍光輝度の低下が見られた モネンシン区においては Rumen bacterium のバンドの蛍光輝度の向上と Clostridium leptum のバンドの蛍光輝度の低下が認められた 第 Ⅳ 期においては 対照区と比較して 4g 区で Clostridium thermoscinogenes BacteriumYE57 および Clostridium leptum のバンドの蛍光輝度の向上が見られた BLCS8g 区では Quinella ovalis Clostridium leptum および Bacterium YE57 の蛍光輝度の向上が見られ Rumen bacterium の蛍光輝度の低下が認められた モネンシン区においては Clostridiumthermoscinogenes および Swine manure bacterium のバンドの蛍光輝度の向上が見られた
37 4-18 PCR-DGGE 法によるルーメンフローラ解析に関する考察 1) 真正細菌の解析 1) -1 BLCS ルナシータに含まれる細菌 BLCS ルナシータに含まれる細菌として Bacillus 属および Streptococcus 属が同定された Bacillus 属として同定された細菌はアルカリ環境 ( 至適 ph10.5) を好むものであり (Krulwich et al. 1985) ルーメン内は中性環境( 高橋 2007) であることから ルーメン内において活発に増殖した可能性は低いと考えられる しかし ルーメン内に生存し続けて機能を発揮していたと仮定すると Bacillus 属の中にはフマル酸還元酵素を有する種もあるため (Schnorpfeil M et al. 2001) 水素を消費することが可能であり また 酪酸が主要な代謝産物であるため その生成過程においても水素を消費することができると考えられる この様な水素の消費により メタン生成菌が利用する基質を減少させ メタン生成菌数を低下させうると考えられる また BLCS ルナシータ投与区において酪酸濃度が上昇しているが ( 高橋 2007) その一部の生産を当該 Bacillus 属が担っていた可能性も考えられる Streptococcus 属として同定された細菌は低 ph 環境下 (5.0 以下 ) で乳酸を産生するが 本試験に用いた羊のルーメン内 ph は中性 ( 高橋 2007) であるため 乳酸を産生した可能性は低いと考えられる 一方 Streptococcus 属の主要代謝産物はギ酸および酢酸であるが ギ酸の多くは水素と二酸化炭素に分解され その後メタンに変換される BLCS ルナシータ 4g 区および 8g 区においてメタン生成が抑制される傾向を示したこと ( 高橋 2007) およびルーメン内の酢酸濃度が高かったこと ( 高橋 2007) から BLCS ルナシータに含まれる Streptococcus 属がルーメン内で生存していた場合 ギ酸よりも酢酸が主要な代謝産物になったと考えられる しかし Bacillus 属および Streptococcus 属のいずれも BLCS ルナシータ投与区では DGGE のバンドとして検出されなかった このため これらの細菌がルーメン内で中心となって VFA 産生に貢献していた可能性は低いと考えられる 1) -2 BLCS ルナシータ投与によるルーメンフローラの変化 BLCS ルナシータ投与 4g および 8g 区においては 第 Ⅰ 期 第 Ⅱ 期 第 Ⅲ 期で Clostridiumthermosuccinogenes または Clostridium sp. FG4 のバンドの蛍光輝度が対照区と比較して向上することが認められ 4g 区においては第 Ⅳ 期においても蛍光輝度の向上が見られた Clostridium thermosuccinogenes と Clostridium sp. FG4 は相同性の数値が同じであり 二つの菌は性質が近いと考えられる Clostridium thermosuccinogenes は多糖類のイヌリンを唯一の炭素源として利用可能であるが (Sridhar et al. 2000) BLCS ルナシータの吸着剤として用いたフスマにイヌリンが含まれていることが報告されている (Roland N et al.1995) このため フスマまたはイヌリンの羊への給与によりこれらの細菌が増加した可能性も考えられる しかし Clostridium thermosuccinogenes が増加した原因が BLCS ルナシータに含まれる細菌か吸着剤による影響なのかは不明である これを明らかにするためには BLCS ルナシータの吸着剤に細菌を添加していないものまたは BLCS ルナシータを滅菌したものを対照区の羊に給与して その時のルーメンフローラを解析することが必要であると考えられる 一方 Clostridium thermosuccinogenes はフマル酸還元酵素を有しており (Drent WJ et
38 al. 1991) この酵素の反応過程で水素を消費することができる その結果 メタン生成菌の基質を減少させ メタン生成菌数を低下させた可能性も考えられる さらに Clostridium thermosuccinogenes の主要代謝産物はコハク酸と酢酸であり (Drent WJ et al. 1991) BLCS ルナシータ投与区においてルーメン内で酢酸濃度が高かった結果 ( 高橋 2007) に対して本菌が寄与した可能性も考えられる Clostridium sp. FG4 の主要代謝産物は酢酸であり Clostridium thermosuccinogenes と高い相同性 (99.8%) を有している (Kato S et al. 2005) そのため Clostridium sp. FG4 はフマル酸還元酵素を有している可能性が高いと考えられ 本菌も水素の消費に関与している可能性が高いと考えられる また BLCS ルナシータ投与 4g 区においては第 Ⅳ 期で 8g 区では第 Ⅰ 期および第 Ⅳ 期で Clostridium leptum が対照区と比較して増加していたことが認められている Clostridium leptum は酪酸産生能を有しており その代謝過程において水素を消費する その結果 メタン生成菌の基質を減少させ メタン生成菌数を低下させた可能性が考えられる また BLCS ルナシータ投与区においては対照区と比較して酪酸濃度が高いこと ( 高橋 2007) の原因となっている可能性も考えられる Succiniclasticum ruminis も対照区と比較して BLCS ルナシータ投与 8g 区において DGGE バンドの蛍光輝度の向上が第 Ⅰ 期および第 Ⅱ 期で見られた 本菌はコハク酸をプロピオン酸に代謝する (van Gylswyk NO 1995) この代謝過程においては水素の授受は関与していないが BLCS ルナシータ投与区において Succiniclasticum ruminis が増加したことは 水素消費に関与するフマル酸還元酵素によるコハク酸のプロピオン酸への代謝が行われていたことを示している可能性がある BLCS ルナシータには吸着剤として米ぬか フスマ 大豆粕などの易分解性タンパク質が使われている ( 高橋 2007) にも関わらず 8g 区でルーメンからの尿中への窒素流出は減少する傾向が示されている ( 高橋 2007) したがって BLCS ルナシータ投与により増加した細菌 (Clostridium thermosuccinogenes Clostridium leptum 増加時期および投与区は前出 ) により菌体タンパク質の合成が促進された可能性が考えられる モネンシン区においては VFA 濃度の低下 アンモニア態窒素濃度の低下 メタン発生量の低下 プロトゾア数の低下が見られている ( 高橋 2007) BLCS ルナシータ投与区とはメタン発生量の低下のみが共通し 他は異なる結果が得られている DGGE の解析結果においては モネンシン区で第 Ⅱ 期および第 Ⅳ 期で Swine manure bacterium が増加し BLCS ルナシータ投与区とは異なるルーメンフローラを構築していることが明らかとなった このため BLCS ルナシータによるメタン発生の低下はモネンシンによるメタン発生の低下とは異なるメカニズムによるものであると考えられる 2) 古細菌の解析羊のルーメン内におけるメタン生成菌をクローンライブラリーにより測定した過去の研究例では メタン生成菌の約 96% が Methanobrevibacter 属であることが報告されている (Wright AD et al. 2004) 本試験においても出現した主な DGGE バンドの同定結果は Methanobrevibacter sp. であったことから 過去の報告と一致するものであった Methanobrevibacter sp. は水素資化性メタン生成菌であることから BLCS ルナシータを
39 投与した羊において 水素の授受に関与する真正細菌の増減が生じたことがルーメン内の メタン発生に密接に関連していたものと考えられる 3) 真核生物の解析 DGGE 法による真核生物の解析対象として 元々ルーメン内に生息しているものと BLCS ルナシータに含まれる酵母が考えられる しかし 本試験の DGGE 解析の結果 全くバンドが認められなかった その原因として次の二点が考えられる その一つは検出感度の問題である ルーメン内には約 104 個 /ml の真核生物が存在しているとされており ( 板橋 2007) PCR-DGGE の検出感度 (103 個 /ml) に近似した数値である BLCS ルナシータには酵母が含まれており 増殖すれば検出されたと考えられるが 検出されなかった結果から 酵母がルーメン内において活発に増殖しなかった可能性がある 対照区と比較して BLCS ルナシータ 4g 投与区において繊維消化率 (NDF ヘミセルロース ) がやや低下する傾向が見られた ( 高橋 2007) が 8g 区においては対照区と同程度に回復していることが認められた ( 高橋 2007) BLCS ルナシータ 4g 区においては吸着剤の影響により繊維消化率が低下したと思われる 酵母発酵培養物 (Saccharomyces cerevisiae) の牛ルーメン液への添加について in vitro 試験を行った過去の報告では VFA 生成とプロピオン酸生成が高まり 繊維分解菌数が増加して繊維消化率が約 10% 高まることが報告されている (Lila et al. 2006) さらに BLCS ルナシータの投与により VFA 生成とプロピオン酸生成が高まる結果が得られている ( 高橋 2007) そこで 8g 区においても繊維消化率が回復した理由として BLCS ルナシータに含まれる酵母がルーメン内において機能していた可能性が考えられる これらを明らかにするためには ルーメン液に含まれる酵母を計数することが必要と考えられる また BLCS ルナシータに用いる細菌の吸着剤として 繊維消化率に影響を与えない物質を検討する必要があると思われる 5. 試験 2. T-RFLP 解析によるルーメンフローラの解析 5-1 T-RFLP 法のフロー 図 5-1 に T-RFLP 法によるルーメンフローラ解析のフローを示す 1. ルーメン液からの DNA 抽出 精製 2. PCR 反応 3. T-RFLP 解析 4. T-RF データを利用した細菌の推定 図 5-1 T-RFLP 法によるルーメンフローラ解析のフロー
40 1) ルーメン液からの DNA 抽出 精製 試験 1 でルーメン液から抽出した DNA 溶液を用いた また Ⅱ 期以降は前の期の試験の 影響を受けている可能性があるため Ⅰ 期の試料のみを用いた 2) PCR 反応試験 1 で抽出した DNA 溶液 (Ⅰ 期のみ ) を用いて PCR を行った 試験に用いたプライマーは FAM-27f :AGAGTTTGATCCTGGCTCAG[5' 末端を 6'-carboxyfluorescein(FAM) で標識 ] および 1492r:GGCTACCTTGTTACGACTT である PCR 増幅反応条件を図 5-2 に示す 94 2min 94 30sec 55 30sec 30cycles 68 90sec 68 7min 4 図 5-2 T-RFLP 解析のための PCR 条件 3) T-RFLP 解析 PCR 増幅産物 5 μ l を T-RFLP 解析において一般的に用いられている制限酵素 AluI(Nippon Gene) HhaI(Nippon Gene) および MspI(Nippon Gene)0.5μl を用いて 37 で 10 時間処理した後 95 で 10 分処理することで酵素を失活させた 失活処理液 1μl サイズスタンダード (Gene Scan 500LIZ, Applied BioSystems 社 )0.3μl HiDi ホルムアミド (Applied BioSystems 社 )12μl を混合し 95 で 2 分処理し DNA を一本鎖にした 得られた反応液を ABI PRISM 310DNA シーケンサを用いて POP-4 polymer ラン ( 変性系電気泳動 ) を行い 制限酵素断片長 ( 以下 T-RF と略す ) の蛍光強度を Gene Scan により読み込んだ Gene Mapper ソフトウェア (Applied BioSystems 社 ) により T-RF の蛍光強度の定量解析を行った 4) T-RF データを利用した細菌の推定試験 1 において EMBL-FASTA 解析により同定した細菌の 16S rdna 配列を Gene Bank から入手し 三種類の制限酵素の切断サイトを検索することにより 同定した細菌の理論上の T-RF を求めた 理論上の T-RF の数値と T-RFLP 解析により得られた T-RF の数値
41 を比較することにより細菌を推定した 5-2 結果表 5-4 に EMBL-FASTA により同定した細菌の 16S rdna を三種類の制限酵素処理した時の理論上の T-RF を示した DDBJ-FASTA と比較して EMBL-FASTA による検索においては相同性が同等もしくはそれ以上であるため 表 5-4 においては EMBL-FASTA の結果を用いた Ⅰ 期の試料を用いた AluI 処理による T-RFLP のチャートを図 5-3 に示し HhaI 処理および MspI 処理による T-RFLP のチャートを図 5-4 図 5-5 にそれぞれ示した また T-RF ピークの高さ 面積を示した表について AluI 処理の結果を表 5-5 に HhaI 処理および MspI 処理による結果について表 5-6 表 5-7 にそれぞれ示した AluI 処理の結果において 対照区と比較して BLCS ルナシータ投与区 (4g 区および 8g 区 ) でピーク高が増加した主な T-RF について述べる なお 括弧内には表 5-4 に示した T-RF と近似する細菌名を示す 増加した主な T-RF は 43~45bp 57bp(Bacterium Km5 8g 区のみ ) 68bp 82bp 87bp(Clostridium leptum 8g 区のみ ) 109bp 110bp 134bp 282bp(8g 区のみ ) 283bp(8g 区のみ ) および 436bp(Quinell ovalis 8g 区のみ ) であった ( 図 5-3 に赤矢印で示す ) 一方 対照区と比較して 4g 区および 8g 区でピーク高が低下した主な T-RF は 70bp のみであった ( 図 5-3 に青矢印で示す ) また BLCS ルナシータの T-RFLP 結果で得られたピークの中で 4g 区または 8g 区においても同じ T-RF のサイズで出現したピークは認められなかった HhaI 処理の結果において 対照区と比較して BLCS 投与区 (4g 区および 8g 区 ) でピーク高が増加した主な T-RF は 67bp(Bacterium Km5 8g 区のみ ) 110bp 126bp 127bp 189bp 209bp(8g 区のみ ) 241bp(8g 区のみ ) 383bp(Rumen bacterium 8g 区のみ ) および 560bp(Clostridium thermosuccinogenes 8g 区のみ ) であった ( 図 5-4 に赤矢印で示す ) 一方 対照区と比較して BLCS 投与区でピーク高が低下した主な T-RF は 571bp であった ( 図 5-4 に青矢印で示す ) MspI 処理の結果については 対照区と比較して BLCS 投与区 (4g 区および 8g 区 ) でピーク高が増加した主な T-RF は 77bp 89bp 307bp(8g 区のみ ) 465bp(Bacterium Km5 8g 区のみ ) であった ( 図 5-5 に赤矢印で示す ) 一方 対照区と比較して BLCS 投与 4g および 8g 区でピーク高が低下した主な T-RF は 52bp 58bp 70bp 95bp 156bp 168bp 169bp 170bp 237bp(Clostridium leptum) 268bp 461bp 481bp 493bp および 495bp であった ( 図 5-5 に青矢印で示す )
42 表 5-4 各制限酵素による切断時の T-RF(EMBL-FASTA の同定結果を使用 )
43
44
45
46
47 図 5-3 AluI 処理による Ⅰ 期試料の T-RFLP 結果
48 図 5-4 HhaI 処理による Ⅰ 期試料の T-RFLP 結果
49 図 5-5 MspI 処理による Ⅰ 期試料の T-RFLP 結果
50 5-3 T-RFLP 解析によるルーメンフローラの解析に関する考察各制限酵素処理において BLCS ルナシータと BLCS ルナシータ投与区で出現したピークの中で同一と思われる T-RF は認められず 投与した BLCS ルナシータに含まれていた細菌がルーメン内で活発に増殖した可能性は低いと考えられ DGGE 解析の結果を裏付けるものとなった また BLCS ルナシータ投与 8g 区においては対照区と比較して Clostridium leptum Clostridium thermosuccinogenes と予測されるピーク高が増加することが認められ DGGE の蛍光輝度の結果と一致するものであった 6. 試験 3. MPN-PCR 法を用いたメタン生成菌の定量 6-1 MPN-PCR 法によるメタン生成菌の定量のフロー 図 6-1 に MPN-PCR 法によるメタン生成菌の定量のフローを示した 1. PCR 反応に関する予備検討 2. MPN-PCR 法 3. 最確数表によるメタン生成菌の定量 図 6-1 MPN-PCR 法によるメタン生成菌の定量 6-2 PCR 反応に関する予備検討ルーメン内に存在する全てのメタン生成菌を定量することを目的として Methanobacteriaceae 目 Methanococcales 目 Methanosarcinales 目 Methanomicrobiales 目および Methanosaeta 属に特異的なプライマーを用いて PCR を行った 表 6-1 にメタン生成菌を定量するために用いたプライマーの配列を示す なお フォワードプライマー (109F) は古細菌に共通した配列であり リバースプライマーは各メタン生成菌に特異的な配列とした 図 6-2 に Methanobacteriaceae 目を増幅させる PCR 条件を示した 各メタン生成菌を PCR で増幅するための最適条件を明らかにするための予備試験を行い Methanococcales 目および Methanosarcinales 目はアニーリング温度を 58 に設定し Methanomicrobiales 目および Methanosaeta 属はアニーリング温度を 56 に設定した なお アニーリング温度以外の PCR 増幅条件は Methanobacteriaceae 目と同一である PCR 反応を行うための鋳型として用いた DNA 溶液は 1) 試験 1 で用いたもの (0.5ml から抽出 ) 2) Ultra clean soil DNA kit(mobio 社製 ) を用いて試料 0.5ml から抽出を行ったもの 3) 試験 1 では 0.5ml の試料から DNA 抽出を行ったが 高感度で検出を試みるために 4ml の試料から DNA 抽出を行ったもの とした また PCR 阻害物質の除去を目的として Power clean DNA clean-up kit(mobio 社製 ) および BSA(5μg/μl) の添加 (0.8μ l/pcr 反応液 40μl) について検討を行った
 をエタノール沈殿により 10μl に濃縮し 全量を 1% アガロースゲル電気泳動に供すことにより増幅産物の確認を行った また 10 倍希釈液でバンドが見られない試料については DNA 抽出液の原液およびエタノール沈殿による 10 倍濃縮液を調製して PCR を行った 6-4 最確数表によるメタン生成菌の定量 MPN-PCR")
51 表 6-1 メタン生成菌の定量に用いたプライマー 94 4min 94 45sec 50 1min 30cycles 72 1min 72 7min 4 図 6-2 Methanobacteriaceae 目を増幅させる PCR 条件 6-3 MPN-PCR 予備検討の結果を受けて DNA 抽出液の 10 倍希釈液 100 倍希釈液および 1000 倍希釈液を用いて各希釈液につき 5 反復の PCR を行い PCR 増幅産物 (40μl) をエタノール沈殿により 10μl に濃縮し 全量を 1% アガロースゲル電気泳動に供すことにより増幅産物の確認を行った また 10 倍希釈液でバンドが見られない試料については DNA 抽出液の原液およびエタノール沈殿による 10 倍濃縮液を調製して PCR を行った 6-4 最確数表によるメタン生成菌の定量 MPN-PCR 法で出現したバンド数に基づいて最確数表からメタン生成菌の定量を行った
 また 図 6-4 の各レーンに流した試料の条件を表 6-2 に示す DNA 抽出キットおよび試料量を 4ml に増やして DNA を抽出した方法では BSA を添加しても PCR 増幅産物は認められなかった しかし 試験 1 で抽出した DNA を鋳型として BSA を添加して PCR")
52 6-5 結果図 6-3 に Methanococcales 目 Methanomicrobiales 目 Methanosarcina 目 Methanosaeta 属および Methanobacteriaceae 目の増幅結果を示す その結果 Methanobacteriaceae 目のみで検出された 図 6-3 メタン生成菌の検出結果 図 6-4 に PCR 阻害物質除去による効果および試料濃縮による影響についての結果を示す ( 図中の M はマーカーを示す ) また 図 6-4 の各レーンに流した試料の条件を表 6-2 に示す DNA 抽出キットおよび試料量を 4ml に増やして DNA を抽出した方法では BSA を添加しても PCR 増幅産物は認められなかった しかし 試験 1 で抽出した DNA を鋳型として BSA を添加して PCR を行うことにより PCR 増幅産物が認められた

53 図 6-4 PCR 阻害物質除去による効果および試料濃縮による影響 表 6-2 図 6-4 に示した電気泳動画像における各レーンに流した試料の条件 表 6-3 に MPN-PCR 法で検出されたバンド数を示す なお 10 倍濃縮液および DNA 抽 出液の原液は PCR 阻害物質が増加することや濃縮時に DNA ロスが生じた可能性を考慮し 参考データとした
54 表 6-3 MPN-PCR で検出されたバンド数 表 6-3 に示したバンド数から最確数表に基づいて細菌数を算出した結果を表 6-4 に示した Ⅰ 期およびⅡ 期においては対照区と比較して全ての試験区でメタン生成菌数の低下が認められた Ⅲ 期においては 4g 区においてメタン生成菌数が対照区と比較してやや多い結果となった Ⅳ 期においては全ての試験区で検出限界以下となった
55 表 6-4 メタン生成菌数に及ぼすモネンシンおよび BLCS ルナシータの効果 1) DNA 抽出液を 10 倍濃縮することにより PCR 阻害物質が増加することや濃縮時に DNA ロスが生じる可能性を考慮し 10 倍希釈以降のデータを使用して定量値を算出した 6-6 MPN-PCR 法を用いたメタン生成菌の定量に関する考察牛ルーメン内においてメタン生成菌は約 105~106 個 /ml 存在することが報告されているが ( 小原 2006) 羊を用いた本試験においては約 104~105 個 /ml と数値が近似していた 反芻動物である牛と羊のルーメン内には同程度の密度でメタン生成菌が生息していると考えられることから 今回用いた DNA 抽出法および PCR 条件はメタン生成菌の定量に適したものであると考えられる BLCS ルナシータ投与区においては Ⅰ 期およびⅡ 期において対照区よりもメタン生成菌数の低下が見られており BLCS ルナシータの効果によるものである可能性が考えられた BLCS ルナシータの 8g 区ではⅡ 期以降メタン生成菌数は検出限界以下となり より強力にメタン生成菌数を低減させた可能性が示唆された 一方 4g 区のⅢ 期においては対照区よりもメタン生成菌数が多いという結果となった BLCS ルナシータ 4g 投与では効果が弱かったことやⅡ 期試験の影響が残っていた可能性もあるかもしれないと考えられる Ⅳ 期においては 全ての区においてメタン生成菌数は検出限界以下になっていた 同様にⅢ 期試験の影響が残っていたことも可能性の一つとしてあるのかもしれない メタン生成菌数の全ての試験区の平均値は第 Ⅰ 期より第 Ⅱ 期が少なく 第 Ⅱ 期よりも第 Ⅲ 期が少なく 第 Ⅲ 期よりも第 Ⅳ 期が少ない結果となっている このため 試験間隔を決めるための検討を行うことが今後の課題と考えられる また BLCS ルナシータ 8g 区においては )-2 に示した通り 酪酸を産生する Clostridium leptum が増加することが明らかとなった 高橋らの実験 (2007) では BLCS ルナシータ 8g 区においてルーメン内の酪酸濃度が上昇し それに対応するようにルーメン内 ph が低下している ( 高橋 2007) ことが示されている メタン発生量は ph が 0.1 低下すると 10% 程度低下することが示されているが (Russell JB 1998) BLCS ルナシータ 8g 区において ルーメン内 ph の経時的変化を測定した結果 対照区と比較してルーメン液採取後 4 時間から 8 時間では ph が 0.1 程度低下したことが示されている ( 高橋 2007) ルーメン内 ph が低下することにより メタン生成菌またはメタン生成菌が付着するプロトゾアの数が 8g 区において活動低下または死滅し ルーメンからのメタン発生量が低下した可能性が考えられる すなわち BLCS ルナシータの投与で 酪酸を産生する Clostridium
56 leptum が増加して ph が低下し メタン生成菌が死滅してメタン発生が低下した可能性が考えられる モネンシン区と 4g 区においては 対照区と比較してメタン生成菌数の低下はわずかなものであり メタン発生量の低下 ( 高橋 2007) と一致するものであると考えられる 一方 8g 区においては対照区と比較してメタン生成菌数は 1/10 以下に低下していることが示されたが メタン発生量は対照区と比較して 5~10% 低減するに留まっていた ( 高橋 2007) BLCS ルナシータに含まれる物質が PCR を阻害して BLCS ルナシータ 8g 投与後のルーメン内のメタン生成菌数の検出に影響を与えていたのではないかと考えられる モネンシン区においてはプロトゾア数の低減が認められ ( 高橋 2007) プロトゾアは繊維分解活性を有することから繊維消化率の低下が予想されたが 中性デタージェント繊維 (NDF) 酸性デタージェント繊維 (ADF) の消化率は対照区と同じ程度であった ( 高橋 2007) これに対してはプロトゾア数の低下が繊維消化率に及ぼす影響が小さかったことと 繊維分解菌数が増加したことの 2 つの可能性が考えられる 7まとめ BLCS ルナシータに含まれる細菌は投与区においてルーメン液から検出されず ルーメン内において中心的に機能している可能性は低いことが示された しかし 他の細菌の活性化を通して機能を発揮している可能性は残されている BLCS ルナシータ投与区においては Clostridium thermosuccinogenes Clostridium leptum Succiniclasticum ruminis が増加しており いずれの細菌も水素を消費する代謝経路を有することから メタン発生の低減と関係している可能性があると考えられる BLCS 投与区においては多くの場合 対照区と比較してメタン生成菌数の低下が認められた 8. 提言ルーメンフローラの変化が BLCS ルナシータに含まれる細菌か吸着剤による影響であるのか明らかにするため BLCS ルナシータの吸着剤に細菌を添加していないもの または BLCS ルナシータを滅菌したものを対照区の羊に給与する必要があると考えられる BLCS ルナシータに含まれる酵母がルーメン内において機能しているか明らかにするため 培養法により計数することや繊維分解菌を MPN-PCR 法等の手法を用いて定量することが必要と考えられる 本試験で生息が確認された Methanobrevibacter は培地中の増殖速度が真正細菌の数十分の 1 程度と遅く ルーメン内における増殖速度は明らかにされていないことから 試験間隔を決めるために BLCS ルナシータおよびモネンシン投与後のメタン生成菌数を経時的に測定することが必要であると考えられる また ルーメンからのメタン発生は粗飼料多給により低下することが報告されている (Russell JB 1998) ことから 給与飼料の違いがメタン生成菌数に及ぼす影響を明らかにし メタン生成菌数が高まる飼料を用いて試験を行
57 うことが必要であると思われる BLCS ルナシータ 4g 区において繊維消化率が低下して いることから BLCS ルナシータの吸着剤として繊維消化率に影響を与えない物質を検討 する必要も考えられる
58 9. 参考資料 Drent WJ, Lahpor GA, Wiegant WM, Gottschal JC (1991) Appl EnvironMicrobiol (1991) 57: 板橋久雄 (2007) ルーメン研究会報 18: Kato S, S Haruta, ZJ Cui, M Ishii, Y Igarashi(2005) Appl Environ Microbiol. 71: Krulwich TA, Aqus R, Schneier M, Guffanti AA (1985) J Bacteriol. 162: Lila ZA, N Mohammed, T Takahashi, M Tabata, T Yasui, M Kurihara, S Kanda and H Itabashi (2006) Anim Sci J. 77: 小原 (2006) ルミノロジーの基礎と応用. 農文協. Roland N, Nugon-Baudon L, Andrieux C, Szylit O (1995) Br J Nutr. 74: Russell JB (1998) J Dairy Sci. 81: Schnorpfeil M, Janausch IG, Biel S, Kroger A, Unden G (2001) Eur J Biochem. 268: Sridhar J, Eiteman MA, Wiegel JW (2002) Appl Environ Microbiol. 66: 高浦一希ら (2006) 第 106 回日本畜産学会大会高橋潤一 (2007) 生菌剤 (BLCS ルナシータ ) がめん羊のルーメンメタン生成 飼料消化率及び窒素 エネルギー出納に及ぼす影響. Van Gylswyk NO (1995) Int J Syst Bacteriol. 45: Wright AD, Williams AJ, Winter B, Christophersen CT, Rodgers SL, Smith KD (2004) Appl Environ Microbiol. 70: Yoshida N, Takahashi N, Hiraishi A (2005) Appl Environ Microbiol. 71:
59 2) 乳酸菌の産生産生するするプロテアーゼプロテアーゼ耐性抗菌性物質 (PRA) の影響 1 目的味の素株式会社殿においては ルーメンから発生する温室効果ガスであるメタンを抑制する効果の見られる乳酸菌由来の抗菌物質の開発に取り組まれており 試験管レベルでメタンをほぼ完全に抑制する結果を得ている 抗菌物質をルーメン液に添加した際に結果として増加する細菌は メタン生成菌の基質である水素の利用またはメタン生成菌に対する抗菌効果を有している可能性が考えられる このため この様な機能を持つ細菌はメタン抑制効果を有するプロバイオティクスとしての開発が期待できる 以上より本試験は 抗菌物質をルーメン液に添加した時の細菌相の変化を明らかにすることを目的として行った 2 試料ルーメン液をホルスタイン種から採取し 緩衝液および基質を添加したもの ( 対照区 ) および対照区に乳酸菌培養上清を 10% 添加したもの ( 添加区 ) を それぞれ 24 時間培養し 微生物解析用試料とした また メタン抑制効果を有するモネンシン投与時の細菌相と比較するため チェビオット種去勢羊に給与飼料のみを投与したもの ( コントロール区 ) および 1 日当たりモネンシン 20mg を 2 週間投与したもの ( モネンシン区 ) から採取したルーメン液も微生物解析用試料として用いた
60 3 方法 3-1 PCR- 変性グラジエントゲル電気泳動 (DGGE) 法 図 3--1 に PCR-DGGE 法による微生物群集構造解析のフローを示す 1. 試料からの DNA 抽出 2. PCR 反応 1) 真正細菌を対象とした PCR 反応 (1 回目 ) 2) 真正細菌 PCR 反応 (GC クランプ付き ) 3. アガロース電気泳動による 16S rdna の増幅の確認 4. DGGE 解析 5. DNA バンド切り出しと DNA 抽出 精製 6. PCR 反応 ( プライマー :GC クランプの除去 ) 7. PCR 産物の精製 8. シーケンス解析 10. ホモロジー解析 ( 微生物種の同定 ) DNA バンドが単一にな るまで数回繰り返す 図 3-1 PCR-DGGE 法による微生物群集構造解析のフロー 3-2 試料からの DNA 抽出ルーメン液には DNA 抽出が困難な細菌および PCR 阻害物質が存在すると考えられる このため 本試験では強固な細胞壁を備えた細菌の物理的破壊工程 腐植物質 ( あるいは前駆物質であるリグニン ) の除去工程を含む方法 (Yoshida N et al. 2005) を用いた
61 3-3 PCR 反応 表 3--1 にプライマーの配列を示した 表 3--1 真正細菌を対象とした PCR に用いたプライマーの配列 3-4 真正細菌を対象とした PCR 反応 (1 回目 ) 抽出した DNA を鋳型として 真正細菌用のプライマー (341F 907R) を用いて 16SrDNA を増幅した PCR 条件を図 3-2 に示した PCR 反応 (Primer : 341F, 907R) 96 5min 96 1min 55 30sec 25cycle 72 1min 72 5min 4 図 3-2 真正細菌の 16SDNA を標的とした PCR 反応条件 3-5 真正細菌を対象とした PCR 反応 (GC クランプ付き ) DGGE ゲルを用いた電気泳動を行うため GC クランプ付 (GC に富む配列 ) プライマーを用いた PCR を行った 具体的には図 3-2 で得られた PCR 産物を鋳型として 真正細菌用 GC クランプ付きプライマー (341FGC,534R) を用いて PCR を行い ( 図 3-3) アガロースゲル電気泳動にて PCR 産物の増幅を確認した
62 PCR 反応 (Primer : 341FGC, 534R) 96 5min 96 1min 55 30sec 25cycle 72 30sec 72 5min 4 図 3-3 真正細菌の 16SDNA を標的とした PCR 反応条件 (GC クランプ付 ) 3-6 DGGE 解析広範囲のバンドが得られるように ゲル中の DNA 変性剤の濃度勾配を 30-70%( 変性剤 100% とは 7M 尿素 40% ホルムアミドに相当 ) として DGGE 解析を行った また 重合剤の濃度は 低濃度に設定し ゲルを 4 時間固化させることにより グラジエントの精度を高めたゲルを作成した さらに バンドの分離能を高めるため 電圧を低くし 泳動時間を長く設定した ( 表 3-2) 表 3-2 DGGE ゲル濃度と泳動条件 ポリアクリルアミドゲル濃度 :10% 変性剤濃度勾配 :30-70% 重合剤濃度 10% Ammonium Persulfate :0.7% TEMED :0.07% 泳動電圧 :70V 泳動時間 :960min 泳動温度 : DNA バンドの切り出しと DNA 抽出 精製電気泳動後 エチジウムブロマイドで染色を行い 脱染色後 UV トランスイルミネーター上でバンドを検出した 検出したバンドを滅菌されたメスで切り出し 1.5ml チューブに入れた 切り出したゲルに 100μl の TE buffer(ph8.0) を加え 室温で一晩振とうさせた後 エタノール沈殿法により DNA の抽出 精製を行った なお 分離能が低く PCR 複数のバンドが混在している可能性のあるバンドについては バンドが単一になるまで GC
63 クランプ付 PCR および DGGE 解析を繰り返した 3-8 PCR 反応 (GC クランプの除去 ) 各バンドから抽出した DNA を鋳型として シーケンス反応を行うため GC クランプを 外す PCR を行った 1) GC クランプ無し PCR GC クランプを外す PCR の条件を図 3-4 に示した PCR 反応 (Primer : 341F, 534R) 96 5min 96 1min 55 30sec 25cycle 72 30sec 72 5min 4 図 3-4 GC クランプの除去に用いた PCR 反応条件 3-9 PCR 産物の精製アガロースゲル電気泳動により PCR 産物を確認し 次いで アガロースから目的バンドを切り出して 精製キット (MonoFas ; ジーエルサイエンス社製 ) により PCR 産物を精製した 3-10 シーケンス解析 3-9 で精製を行った PCR 産物を鋳型として ダイターミネーター法 (Big Dye Terminator v1.1 Cycle Sequencing Kit;ABI 社製 ) によりシーケンシング反応を行った シーケンス解析は ABI Prism310 Genetic Analyzer(ABI 社製 ) を用いて行った 3-11 ホモロジー解析 ( 微生物種の同定 ) 解読した各バンドの DNA シーケンスを基に インターネットを利用した FASTA 検索によ り DDBJ のデータベースから微生物種の同定を行った
 対照 : 対照区 添加 : 添加区 ( ホルスタイン種を用いた試験 ) 図 4-1 DGGE 解析結果 (")
64 4. 結果 DGGE バンドパターンを図 4-1 に示した また 塩基配列を決定したバンドを矢印で示し 最も高い相同性を示した細菌名を示した なお 出現した全ての細菌はルーメンからの分離例が報告されている con: コントロール区 mo: モネンシン区 ( チェビオット種去勢羊を用いた試験 ) 対照 : 対照区 添加 : 添加区 ( ホルスタイン種を用いた試験 ) 図 4-1 DGGE 解析結果 ( 白黒反転画像 )

65 4-1 シーケンス解析結果 シーケンス解析の結果決定した塩基配列を表 4-2 に示した 表 4-2 決定した塩基配列
66 4-2 ホモロジー解析結果 ( 微生物種の同定 ) 各バンドの塩基配列に基づいて DDBJ-FASTA 検索を行い ホモロジーの高かった上位 5 種を表 4-3 に示した また それらの Identity( 相同性 ; % で表す ) と Overlap した領域 (nt; 塩基対数 ) を表記した 表 4-3 同定結果
67
68
69 4-3 同定した細菌の生態的特徴 表 4-4 に同定した細菌の生態的特徴を示した 表 4-4 同定した細菌の生態的特徴
70
71
72
73
74
75 5 考察 1) 抗菌物質添加時のルーメンフローラ解析における DGGE の有用性 PCR-DGGE 法による解析により 抗菌物質添加時に無添加と比較してルーメン内で増減する細菌を捉えることができた 表 5-1 に抗菌物質添加時に増加した細菌 減少した細菌 変化しなかった細菌について 最も相同性の高い細菌名およびグラム陽性 陰性の区別を記載した 表 5-1 抗菌物質添加時に増加した細菌 減少した細菌 変化しなかった細菌およびグラム 陽性 陰性の区別 増加した細菌 無添加と比較して DGGE バンドの蛍光輝度の上昇またはバンドの出現 ( 目視観察による ) 減少した細菌 無添加と比較して DGGE バンドの蛍光輝度の低下またはバンドの消失 ( 目視観察による ) 変化しなかった細菌 無添加と比較して DGGE バンドの蛍光輝度が変化しない ( 目視観察による )
76 2)-1 抗菌物質添加により増加した細菌によるメタン抑制について抗菌物質投与時に増加する細菌は メタン生成菌の基質である水素の利用またはメタン生成菌に対する抗菌効果等を有している可能性が考えられる このため 表 5-1 に示した抗菌物質添加により増加した細菌の中に メタン抑制に関与しているものが存在する可能性がある 表 5-1 に示した抗菌物質添加時に増加した細菌の中で メタン抑制に関与している細菌が存在すると仮定した場合 Streptococcus sp. および Butyrivibrio sp. の 2 種類が最も関与している可能性が高いと推察される ルーメン内におけるこれまでの分離例 (Tajima K et al. 2001b) から推察すると これらの細菌の属種はそれぞれ Streptococcus bovis および Butyrivibrio fibrisolvens であると思われる この仮定が正しい場合のメタン抑制メカニズムの仮説を以下に示す 1Streptococcus bovis によるメタン抑制メカニズム ( 仮説 ) Streptococcus bovis LRC0476 HC5 株はバクテリオシンを産生し HC5 株の産生するバクテリオシンである bovicin HC5 の抗菌メカニズムは不明であるが in vitro で混合ルーメン液からのメタン産生を約 50% 抑制することが報告されている (Lee et al. 2002) bovicin HC5 はグラム陽性菌に対して抗菌作用を有することが報告されている (Houlihan AJ et al. 2006) ため ルーメン内の主なメタン生成菌であるグラム陽性のメタン生成菌 (Methanobrevibacter ruminantium) に対して直接抗菌効果を有している可能性が考えられる しかし DGGE の結果からは真正細菌のグラム陽性菌の減少は見られなかったことから メタン生成菌に対する抗菌効果ではなく メタン生成に関与する酵素活性を抑制している可能性も考えられる また 本菌がバクテリオシンを産生していない株であるために メタン抑制とは全く関係が無い可能性もある 2Butyrivibrio fibrisolvens によるメタン抑制メカニズム ( 仮説 ) 本菌が産生するバクテリオシンは グラム陽性菌に対して抗菌効果を有している (Jennifer et al 2002) ルーメン内に存在する主なメタン生成菌である Methanobrevibacter ruminantium はグラム陽性菌であり このメタン生成菌に対して抗菌効果を示した可能性もある しかし Streptococcus bovis と同様に DGGE の結果からは真正細菌のグラム陽性菌の減少は見られていないため メタン生成に関与する酵素活性を低下させる可能性も考えられる 一方 本菌は不飽和脂肪酸に対する水素添加能を有しており メタン生成菌の基質である水素を消費するが 水素添加のために消費される水素はルーメン内の水素の 2% 以内である (Czerkawski. 1986) このため 不飽和脂肪酸に対する水素添加は 水素の競合によるメタン発生の抑制に対する影響は極めて少ないと考えられる 本菌もバクテリオシンを産生しない株である可能性はあるものの メタン抑制に関与している場合は メタン生成に関与する酵素活性の抑制やバクテリオシンによるメタン生成菌に対する抗菌効果である可能性が高いと推察される 2)-2 メタン抑制効果を目指したプロバイオティクスの開発とその留意点について今後の検討により Streptococcus bovis や Butyrivibrio fibrisolvens がメタン抑制に関与している可能性が高いと結論される場合 全ての株にメタン生成酵素の阻害能やバクテリ
77 オシン産生能があるわけではないと考えられるため 細菌の選抜時に注意を要する Streptococcus bovis の場合 多量に乳酸を産生することから ルーメン内 ph の低下が生じ 病的状態であるルーメンアシドーシスを引き起こす危険性があることが指摘されている (Russell and Hino. 1985) このため 抗菌物質の投与または Streptococcus bovis をプロバイオティクスとして使用する場合には投与量に注意を払う必要があると考えられる また ルーメン内乳酸の 74% が Megasphaera elsdenii により利用されるという報告があり (Counotte et al. 1981) Megasphaera elsdenii をルーメン液に添加した in vitro の試験では 乳酸蓄積量の低下と ph 低下の防止が見られている (Kung and Hession. 1995) このため Streptococcus bovis をプロバイオティクスとして使用する場合は Megasphaera elsdenii 等の乳酸蓄積量を低下させる細菌との併用投与も検討する必要があると思われる 3)-1 抗菌物質の抗菌作用について表 5-1 に示した通り 抗菌物質添加により増加した細菌は全てグラム陽性菌であり 抗菌物質添加により減少した細菌は全てグラム陰性菌であった この結果から 抗菌物質はグラム陰性菌に特異的に抗菌効果を有している可能性が高いと考えられる グラム陰性菌に対する有効な抗生物質としてテトラサイクリン系があり この抗生物質の抗菌効果はタンパク質合成を阻害することによるものである また テトラサイクリン系抗生物質は本試験の結果でも減少の見られた Prevotella 属に対する抗菌作用を有しており 一つの可能性として 本試験で用いた抗菌物質はタンパク質合成を阻害していることが考えられる 一方 抗菌物質添加時に対照区と比較してバンドの蛍光輝度に変化の見られなかったバンド No.7 の Bacterium L4M2 1-7 はグラム染色の結果は不明である しかし バンド No.7 のホモロジー解析の結果では ホモロジーの高かった上位 5 種の中で本菌を除く全てがグラム陰性菌であった このため Bacterium L4M2 1-7 はグラム陰性菌である可能性が高いと推察されるが 本菌は抗菌物質による影響をほとんど受けていないことから 抗菌効果がグラム陰性菌に対するものでは無い可能性もあると思われる 3)-2 抗菌物質添加によるルーメン内揮発性脂肪酸 (VFA) の変化について味の素 殿の試験結果から 抗菌物質をルーメン液に添加して培養するとプロピオン酸濃度が低下する場合が多いとの結果が得られている ( 味の素 年. 未発表 ) プロピオン酸産生に関与している Prevotella 属細菌や優占種ではないもののプロピオン酸を産生するグラム陰性菌が抗菌物質添加による影響で減少することにより プロピオン酸濃度が低下する可能性が考えられる 3)-3 抗菌物質とモネンシンによる抗菌作用の比較モネンシンはメタン抑制効果を有していることから 抗菌物質添加時に増減する細菌はモネンシン投与時に増減する細菌と類似している可能性を考えていたが DGGE の結果からは 同じ挙動を示すバンドは見られなかった 逆に 抗菌物質添加で増加の見られたバンド No.3(Prevotella sp.) はモネンシン投与時には減少が見られた また 抗菌物質添加で減少の見られたバンド No.8(Succinivibrio dextrinosolvens) および No.12(Schwartzia
78 succinivorans) はモネンシン投与時には増加が見られた この様な結果が得られた理由とし て モネンシンが主にグラム陽性菌に対して抗菌効果を有しているのに対して 本試験の 対象とした抗菌物質は 主にグラム陰性菌に対して抗菌効果を及ぼしたことが考えられる 3)-4 抗菌物質がメタン生成菌に及ぼす影響についての仮説本試験は真正細菌の解析のみを行っており メタン生成菌を含む古細菌の解析を実施していないため 正確性に欠けると思われるが メタン生成菌に対する抗菌物質の作用として以下の仮説が考えられる ルーメン内には主要なメタン生成菌であるグラム陽性の Methanobrevibacter ruminantium(whitford et al. 2001) およびグラム陰性の Methanomicrobium mobile (Tajima et al. 2001a) が存在することが知られている 抗菌物質添加により メタン発生はほぼ完全に抑制されている ( 味の素 未発表 ) ことから 抗菌物質は直接的 間接的にグラム陽性 陰性両方のメタン生成菌に対して抗菌効果を有している可能性が考えられる 間接的なメタン生成菌に対する影響として 抗菌物質添加により増加した細菌の中に メタン生成菌との基質 ( 水素 ) の競合や 抗菌物質の産生およびメタン生成に関与する酵素の阻害物質の産生に関与しているものが含まれる可能性が考えられる 4) まとめ 表 5-2 に考察のまとめを示す 表 5-2 考察まとめ 抗菌物質は真正細菌のグラム陰性菌に対して抗菌効果を有している可能性が高い 抗菌物質によるメタン抑制のメカニズムはモネンシンとは異なると推察される 抗菌物質による抗菌効果だけでなく 抗菌物質添加により増加した Streptococcus 属または Butyrivibrio 属が産生するバクテリオシンがグラム陽性のメタン生成菌に対して抗菌効果を有している可能性もある Streptococcus bovis は投与によりルーメンアシドーシスを引き起こす危険性があり プロバイオティクスとして使用する場合には投与方法に注意を払う必要があると考えられる グラム陰性菌の中で主に Prevotella 属の減少によりプロピオン酸の低下が生じている可能性がある 6 提言本試験の結果 抗菌物質の添加によるルーメンフローラの変化を捉えることができたため 今後は PCR-DGGE 法を用いて様々な条件下での抗菌物質添加時のルーメンフローラを解析していくことが考えられる
79 7 引用文献 Counotte GH, RA Prins. Vet Res Commun. 5: Czerkawski JW. Milligan LP, WL Grovum A Dobson (Eds.) pp PrenticeHall, Englewood Cliffs, New Jersey. Jennifer L. Rychlik and James B. Russell. Appl Environ Microbiol March; 68(3): Houlihan AJ Russell JB. J Appl Microbiol. 100: Kung L Jr, AO Hession. J Anim Sci. 76: Lee SS, JT Hsu, HC Mantovani, JB Russell. FEMS Microbiol Lett. 217: Russell JB, T Hino. J Dairy Sci. 67: Tajima K, T Nagamine, H Matsui, M Nakamura, RI Aminov. FEMS Microbiol Lett. 200: a. Tajima K, Aminov RI, Nagamine T, Matsui H, Nakamura M, Benno Y. Appl Environ Microbiol. 67: b. Whitford MF, RM Teather, RJ Forster, BMC Microbiol. 1: Yoshida N, Takahashi N, Hiraishi A. Appl Environ Microbiol. 71:
■リアルタイムPCR実践編
 リアルタイム PCR 実践編 - SYBR Green I によるリアルタイム RT-PCR - 1. プライマー設計 (1)Perfect Real Time サポートシステムを利用し 設計済みのものを購入する ヒト マウス ラットの RefSeq 配列の大部分については Perfect Real Time サポートシステムが利用できます 目的の遺伝子を検索して購入してください (2) カスタム設計サービスを利用する
リアルタイム PCR 実践編 - SYBR Green I によるリアルタイム RT-PCR - 1. プライマー設計 (1)Perfect Real Time サポートシステムを利用し 設計済みのものを購入する ヒト マウス ラットの RefSeq 配列の大部分については Perfect Real Time サポートシステムが利用できます 目的の遺伝子を検索して購入してください (2) カスタム設計サービスを利用する
手順 ) 1) プライマーの設計 発注変異導入部位がプライマーのほぼ中央になるようにする 可能であれば 制限酵素サイトができるようにすると確認が容易になる プライマーは 25-45mer で TM 値が 78 以上になるようにする Tm= (%GC)-675/N-%mismatch
 1) プライマーの設計 発注変異導入部位がプライマーのほぼ中央になるようにする 可能であれば 制限酵素サイトができるようにすると確認が容易になる プライマーは 25-45mer で TM 値が 78 以上になるようにする Tm= (%GC)-675/N-%mismatch") Mutagenesis 目的 ) 既存の遺伝子に PCR を利用して変異を導入する 1 点変異導入方法 ) Quik Change Site-Directed Mutagenesis Kit(Stratagene) のプロトコールを流用 http://www.stratagene.com/products/showproduct.aspx?pid=131 Kit 中では DNA polymerase
Mutagenesis 目的 ) 既存の遺伝子に PCR を利用して変異を導入する 1 点変異導入方法 ) Quik Change Site-Directed Mutagenesis Kit(Stratagene) のプロトコールを流用 http://www.stratagene.com/products/showproduct.aspx?pid=131 Kit 中では DNA polymerase
DNA/RNA調製法 実験ガイド
 DNA/RNA 調製法実験ガイド PCR の鋳型となる DNA を調製するにはいくつかの方法があり 検体の種類や実験目的に応じて適切な方法を選択します この文書では これらの方法について実際の操作方法を具体的に解説します また RNA 調製の際の注意事項や RNA 調製用のキット等をご紹介します - 目次 - 1 実験に必要なもの 2 コロニーからの DNA 調製 3 増菌培養液からの DNA 調製
DNA/RNA 調製法実験ガイド PCR の鋳型となる DNA を調製するにはいくつかの方法があり 検体の種類や実験目的に応じて適切な方法を選択します この文書では これらの方法について実際の操作方法を具体的に解説します また RNA 調製の際の注意事項や RNA 調製用のキット等をご紹介します - 目次 - 1 実験に必要なもの 2 コロニーからの DNA 調製 3 増菌培養液からの DNA 調製
Microsoft Word - FMB_Text(PCR) _ver3.doc
 _ver3.doc") 2.PCR 法による DNA の増幅 現代の分子生物学において その進歩に最も貢献した実験法の1つが PCR(Polymerase chain reaction) 法である PCR 法は極めて微量の DNA サンプルから特定の DNA 断片を短時間に大量に増幅することができる方法であり 多大な時間と労力を要した遺伝子クローニングを過去のものとしてしまった また その操作の簡便さから 現在では基礎研究のみならず臨床遺伝子診断から食品衛生検査
2.PCR 法による DNA の増幅 現代の分子生物学において その進歩に最も貢献した実験法の1つが PCR(Polymerase chain reaction) 法である PCR 法は極めて微量の DNA サンプルから特定の DNA 断片を短時間に大量に増幅することができる方法であり 多大な時間と労力を要した遺伝子クローニングを過去のものとしてしまった また その操作の簡便さから 現在では基礎研究のみならず臨床遺伝子診断から食品衛生検査
Bacterial 16S rDNA PCR Kit
 研究用 Bacterial 16S rdna PCR Kit 説明書 v201802da 微生物の同定は 形態的特徴 生理 生化学的性状 化学分類学的性状などを利用して行われますが これらの方法では同定までに時間を要します また 同定が困難な場合や正しい結果が得られない場合もあります 近年 微生物同定にも分子生物学を利用した方法が採用されるようになり 微生物の持つ DNA を対象として解析を行う方法が活用されています
研究用 Bacterial 16S rdna PCR Kit 説明書 v201802da 微生物の同定は 形態的特徴 生理 生化学的性状 化学分類学的性状などを利用して行われますが これらの方法では同定までに時間を要します また 同定が困難な場合や正しい結果が得られない場合もあります 近年 微生物同定にも分子生物学を利用した方法が採用されるようになり 微生物の持つ DNA を対象として解析を行う方法が活用されています
遺伝子検査の基礎知識
 リアルタイム PCR( インターカレーター法 ) 実験ガイドこの文書では インターカレーター法 (TB Green 検出 ) によるリアルタイム PCR について 蛍光検出の原理や実験操作の流れなどを解説します 実際の実験操作の詳細については 各製品の取扱説明書をご参照ください - 目次 - 1 蛍光検出の原理 2 実験に必要なもの 3 実験操作法 4 結果の解析 1 1 蛍光検出の原理 インターカレーターによる蛍光検出の原理
リアルタイム PCR( インターカレーター法 ) 実験ガイドこの文書では インターカレーター法 (TB Green 検出 ) によるリアルタイム PCR について 蛍光検出の原理や実験操作の流れなどを解説します 実際の実験操作の詳細については 各製品の取扱説明書をご参照ください - 目次 - 1 蛍光検出の原理 2 実験に必要なもの 3 実験操作法 4 結果の解析 1 1 蛍光検出の原理 インターカレーターによる蛍光検出の原理
無細胞タンパク質合成試薬キット Transdirect insect cell
 発現プラスミドの構築 1. インサート DNA の調製開始コドンは出来るだけ 5'UTR に近い位置に挿入して下さい 経験的に ptd1 の EcoRV/KpnI サイトへのライゲーション効率が最も高いことを確認しています 本プロトコルに従うと インサートサイズにも依りますが 90% 以上のコロニーがインサートの挿入されたクローンとして得られます 可能な限り EcoRV/KpnI サイトへ挿入されるお奨めします
発現プラスミドの構築 1. インサート DNA の調製開始コドンは出来るだけ 5'UTR に近い位置に挿入して下さい 経験的に ptd1 の EcoRV/KpnI サイトへのライゲーション効率が最も高いことを確認しています 本プロトコルに従うと インサートサイズにも依りますが 90% 以上のコロニーがインサートの挿入されたクローンとして得られます 可能な限り EcoRV/KpnI サイトへ挿入されるお奨めします
cDNA cloning by PCR
 cdna cloning/subcloning by PCR 1. 概要 2014. 4 ver.1 by A. Goto & K. Takeda 一般的に 細胞または組織由来の RNA から作製した cdna(cdna pool) から 特定の cdna をベクターに組み込む操作を cdna cloning と呼ぶ その際 制限酵素認識配列を付与したオリゴ DNA primer を用いた PCR
cdna cloning/subcloning by PCR 1. 概要 2014. 4 ver.1 by A. Goto & K. Takeda 一般的に 細胞または組織由来の RNA から作製した cdna(cdna pool) から 特定の cdna をベクターに組み込む操作を cdna cloning と呼ぶ その際 制限酵素認識配列を付与したオリゴ DNA primer を用いた PCR
Bacterial 16S rDNA PCR Kit
 研究用 Bacterial 16S rdna PCR Kit 説明書 v201307da 微生物の同定は 形態的特徴 生理 生化学的性状 化学分類学的性状などを利用して行われますが これらの方法では同定までに時間を要します また 同定が困難な場合や正しい結果が得られない場合もあります 近年 微生物同定にも分子生物学を利用した方法が採用されるようになり 微生物の持つ DNA を対象として解析を行う方法が活用されています
研究用 Bacterial 16S rdna PCR Kit 説明書 v201307da 微生物の同定は 形態的特徴 生理 生化学的性状 化学分類学的性状などを利用して行われますが これらの方法では同定までに時間を要します また 同定が困難な場合や正しい結果が得られない場合もあります 近年 微生物同定にも分子生物学を利用した方法が採用されるようになり 微生物の持つ DNA を対象として解析を行う方法が活用されています
大学院博士課程共通科目ベーシックプログラム
 平成 30 年度医科学専攻共通科目 共通基礎科目実習 ( 旧コア実習 ) 概要 1 ). 大学院生が所属する教育研究分野における実習により単位認定可能な実習項目 ( コア実習項目 ) 1. 組換え DNA 技術実習 2. 生体物質の調製と解析実習 3. 薬理学実習 4. ウイルス学実習 5. 免疫学実習 6. 顕微鏡試料作成法実習 7. ゲノム医学実習 8. 共焦点レーザー顕微鏡実習 2 ). 実習を担当する教育研究分野においてのみ単位認定可能な実習項目
平成 30 年度医科学専攻共通科目 共通基礎科目実習 ( 旧コア実習 ) 概要 1 ). 大学院生が所属する教育研究分野における実習により単位認定可能な実習項目 ( コア実習項目 ) 1. 組換え DNA 技術実習 2. 生体物質の調製と解析実習 3. 薬理学実習 4. ウイルス学実習 5. 免疫学実習 6. 顕微鏡試料作成法実習 7. ゲノム医学実習 8. 共焦点レーザー顕微鏡実習 2 ). 実習を担当する教育研究分野においてのみ単位認定可能な実習項目
16S (V3-V4) Metagenomic Library Construction Kit for NGS
 Metagenomic Library Construction Kit for NGS") 研究用 16S (V3-V4) Metagenomic Library Construction Kit for NGS 説明書 v201705da 本製品は イルミナ社 MiSeq で 16S rrna 細菌叢解析を行うための PCR 増幅キットです 細菌 16S rrna 遺伝子の V3-V4 領域を対象とし PCR 酵素に 多様な配列を効率よく増幅可能な Tks Gflex DNA Polymerase
研究用 16S (V3-V4) Metagenomic Library Construction Kit for NGS 説明書 v201705da 本製品は イルミナ社 MiSeq で 16S rrna 細菌叢解析を行うための PCR 増幅キットです 細菌 16S rrna 遺伝子の V3-V4 領域を対象とし PCR 酵素に 多様な配列を効率よく増幅可能な Tks Gflex DNA Polymerase
MightyAmp™ DNA Polymerase Ver.3
 研究用 MightyAmp DNA Polymerase Ver.3 説明書 v201805da MightyAmp DNA Polymerase は 究極の反応性を追求して開発された PCR 酵素であり 通常の PCR 酵素では増幅が困難な PCR 阻害物質を多く含むクルードな生体粗抽出液を用いる場合にも その強力な増幅能により良好な反応性を示します MightyAmp DNA Polymerase
研究用 MightyAmp DNA Polymerase Ver.3 説明書 v201805da MightyAmp DNA Polymerase は 究極の反応性を追求して開発された PCR 酵素であり 通常の PCR 酵素では増幅が困難な PCR 阻害物質を多く含むクルードな生体粗抽出液を用いる場合にも その強力な増幅能により良好な反応性を示します MightyAmp DNA Polymerase
BKL Kit (Blunting Kination Ligation Kit)
") 製品コード 6126/6127 BKL Kit (Blunting Kination Ligation Kit) 説明書 PCR 産物を平滑末端ベクターにクローニングする場合 使用するポリメラーゼ酵素の種類により 3' 末端に余分に付加された塩基を除去し さらに 5' 末端をリン酸化する必要があります 本製品は これらの一連の反応を簡便に短時間に行うためのキットです PCR 産物の末端平滑化とリン酸化を同時に行うことにより
製品コード 6126/6127 BKL Kit (Blunting Kination Ligation Kit) 説明書 PCR 産物を平滑末端ベクターにクローニングする場合 使用するポリメラーゼ酵素の種類により 3' 末端に余分に付加された塩基を除去し さらに 5' 末端をリン酸化する必要があります 本製品は これらの一連の反応を簡便に短時間に行うためのキットです PCR 産物の末端平滑化とリン酸化を同時に行うことにより
豚丹毒 ( アジュバント加 ) 不活化ワクチン ( シード ) 平成 23 年 2 月 8 日 ( 告示第 358 号 ) 新規追加 1 定義シードロット規格に適合した豚丹毒菌の培養菌液を不活化し アルミニウムゲルアジュバントを添加したワクチンである 2 製法 2.1 製造用株 名称豚丹
 不活化ワクチン ( シード ) 平成 23 年 2 月 8 日 ( 告示第 358 号 ) 新規追加 1 定義シードロット規格に適合した豚丹毒菌の培養菌液を不活化し アルミニウムゲルアジュバントを添加したワクチンである 2 製法 2.1 製造用株 名称豚丹") 豚丹毒 ( アジュバント加 ) 不活化ワクチン ( シード ) 平成 23 年 2 月 8 日 ( 告示第 358 号 ) 新規追加 1 定義シードロット規格に適合した豚丹毒菌の培養菌液を不活化し アルミニウムゲルアジュバントを添加したワクチンである 2 製法 2.1 製造用株 2.1.1 名称豚丹毒菌多摩 96 株 ( 血清型 2 型 ) 又はこれと同等と認められた株 2.1.2 性状感受性豚に接種すると
豚丹毒 ( アジュバント加 ) 不活化ワクチン ( シード ) 平成 23 年 2 月 8 日 ( 告示第 358 号 ) 新規追加 1 定義シードロット規格に適合した豚丹毒菌の培養菌液を不活化し アルミニウムゲルアジュバントを添加したワクチンである 2 製法 2.1 製造用株 2.1.1 名称豚丹毒菌多摩 96 株 ( 血清型 2 型 ) 又はこれと同等と認められた株 2.1.2 性状感受性豚に接種すると
Pyrobest ® DNA Polymerase
 Pyrobest DNA Polymerase 説明書 v201102da 対活Pyrobest DNA Polymerase は Pyrococcus sp. 由来の 3' 5' exonuclease 活性 (proof reading 活性 ) を有する耐熱性 α 型 DNA ポリメラーゼです α 型 DNA ポリメラーゼは Pol I 型ポリメラーゼ (Taq DNA ポリメラーゼなど )
Pyrobest DNA Polymerase 説明書 v201102da 対活Pyrobest DNA Polymerase は Pyrococcus sp. 由来の 3' 5' exonuclease 活性 (proof reading 活性 ) を有する耐熱性 α 型 DNA ポリメラーゼです α 型 DNA ポリメラーゼは Pol I 型ポリメラーゼ (Taq DNA ポリメラーゼなど )
スライド 1
 DNA を用いた微生物 分析の現場での活用 平成 18 年 10 月 5 日 B 会場 15:30~15:50 三井農林 ( 株 ) 食品総合研究所微生物分析サービス 衛生管理手法の変化 従来の方法 公定法に基づく微生物検査 一般性菌数 大腸菌群の判定など 実際は HACCP や cgmp の導入 大規模 大量生産 流通の多様化 迅速化 安全性追求意識の高まり より正確正確で迅速迅速かつ簡便簡便な微生物検査方法が必要
DNA を用いた微生物 分析の現場での活用 平成 18 年 10 月 5 日 B 会場 15:30~15:50 三井農林 ( 株 ) 食品総合研究所微生物分析サービス 衛生管理手法の変化 従来の方法 公定法に基づく微生物検査 一般性菌数 大腸菌群の判定など 実際は HACCP や cgmp の導入 大規模 大量生産 流通の多様化 迅速化 安全性追求意識の高まり より正確正確で迅速迅速かつ簡便簡便な微生物検査方法が必要
リアルタイムPCRの基礎知識
 1. リアルタイム PCR の用途リアルタイム PCR 法は 遺伝子発現解析の他に SNPs タイピング 遺伝子組み換え食品の検査 ウイルスや病原菌の検出 導入遺伝子のコピー数の解析などさまざまな用途に応用されている 遺伝子発現解析のような定量解析は まさにリアルタイム PCR の得意とするところであるが プラス / マイナス判定だけの定性的な解析にもその威力を発揮する これは リアルタイム PCR
1. リアルタイム PCR の用途リアルタイム PCR 法は 遺伝子発現解析の他に SNPs タイピング 遺伝子組み換え食品の検査 ウイルスや病原菌の検出 導入遺伝子のコピー数の解析などさまざまな用途に応用されている 遺伝子発現解析のような定量解析は まさにリアルタイム PCR の得意とするところであるが プラス / マイナス判定だけの定性的な解析にもその威力を発揮する これは リアルタイム PCR
組織からのゲノム DNA 抽出キット Tissue Genomic DNA Extraction Mini Kit 目次基本データ 3 キットの内容 3 重要事項 4 操作 4 サンプル別プロトコール 7 トラブルシューティング 9 * 本製品は研究用です *
 組織からのゲノム DNA 抽出キット Tissue Genomic DNA Extraction Mini Kit 目次基本データ 3 キットの内容 3 重要事項 4 操作 4 サンプル別プロトコール 7 トラブルシューティング 9 * 本製品は研究用です * 2 本キットは動物の組織からトータル DNA を迅速に効率よく抽出するようにデザインされています また 細菌 固定組織 酵母用のプロトコールも用意しています
組織からのゲノム DNA 抽出キット Tissue Genomic DNA Extraction Mini Kit 目次基本データ 3 キットの内容 3 重要事項 4 操作 4 サンプル別プロトコール 7 トラブルシューティング 9 * 本製品は研究用です * 2 本キットは動物の組織からトータル DNA を迅速に効率よく抽出するようにデザインされています また 細菌 固定組織 酵母用のプロトコールも用意しています
PrimeSTAR® Mutagenesis Basal Kit
 PrimeSTAR Mutagenesis Basal Kit 説明書 v201107da 目次 I. キットの内容...3 II. 保存...3 III. 本システムの原理...3 IV. プライマー設計について...5 V. 操作...8 VI. 実験例...10 VII. トラブルシューティング...13 VIII. 関連製品...13 IX. 注意...13 2 PrimeSTAR Mutagenesis
PrimeSTAR Mutagenesis Basal Kit 説明書 v201107da 目次 I. キットの内容...3 II. 保存...3 III. 本システムの原理...3 IV. プライマー設計について...5 V. 操作...8 VI. 実験例...10 VII. トラブルシューティング...13 VIII. 関連製品...13 IX. 注意...13 2 PrimeSTAR Mutagenesis
pdf エンドトキシン試験法
 00-0909.pdf.0 エンドトキシン試験法 3 5 6 7 9 0 3 5 6 7 9 0 3 5 6 7 9 30 3 3 33 3 35 36 37 3 39 0 3 5 6 次のように改める. 本試験法は, 三薬局方での調和合意に基づき規定した試験法である. エンドトキシン試験法は, カブトガニ (Limulus polyphemus 又は Tachypleus tridentatus)
00-0909.pdf.0 エンドトキシン試験法 3 5 6 7 9 0 3 5 6 7 9 0 3 5 6 7 9 30 3 3 33 3 35 36 37 3 39 0 3 5 6 次のように改める. 本試験法は, 三薬局方での調和合意に基づき規定した試験法である. エンドトキシン試験法は, カブトガニ (Limulus polyphemus 又は Tachypleus tridentatus)
 4-4 in situ PCR 研究室 in situ PCR 標的菌種と遺伝子 ( 報告年 ) 応用 Dept. of Marine Sciences University of Georgia USA Escherichia coli O157slt II Escherichia coli 16SrDNA Ralstonia eutropha phl (1997,1998,1999,2002)
4-4 in situ PCR 研究室 in situ PCR 標的菌種と遺伝子 ( 報告年 ) 応用 Dept. of Marine Sciences University of Georgia USA Escherichia coli O157slt II Escherichia coli 16SrDNA Ralstonia eutropha phl (1997,1998,1999,2002)
鳥インフルエンザA(H7N9)ウイルス検出マニュアル(第2版)公開用
ウイルス検出マニュアル(第2版)公開用") 鳥インフルエンザ A(H7N9) ウイルス 検出マニュアル ( 第 2 版 ) 1 目次 1 臨床検体またはウイルス培養液からの RNA の抽出 ( 参考 ) ---------- 3 2 リアルタイム RT-PCR(TaqMan Probe 法 ) による鳥インフルエンザ A(H7N9) ウイルスの検出方法 ---------- 4 3 Conventional RT-PCR 法よる鳥インフルエンザ
鳥インフルエンザ A(H7N9) ウイルス 検出マニュアル ( 第 2 版 ) 1 目次 1 臨床検体またはウイルス培養液からの RNA の抽出 ( 参考 ) ---------- 3 2 リアルタイム RT-PCR(TaqMan Probe 法 ) による鳥インフルエンザ A(H7N9) ウイルスの検出方法 ---------- 4 3 Conventional RT-PCR 法よる鳥インフルエンザ
MLPA 法 Q&A 集
 MLPA 法 Q&A 集 目次 1. MLPA 法の導入 Q1. 導入にあたり 何が必要になりますか? Q2. 実験にはどのようなサンプルが必要でしょうか? Q3. サンプルDNAはどれくらい用意すれば良いですか? Q4. キャピラリーシーケンサが自施設に無い場合でも実験はできますか? Q5. FFPE 検体でも実験はできますか? 2. 実験のセットアップ Q1. 実験において重要なポイントはありますか?
MLPA 法 Q&A 集 目次 1. MLPA 法の導入 Q1. 導入にあたり 何が必要になりますか? Q2. 実験にはどのようなサンプルが必要でしょうか? Q3. サンプルDNAはどれくらい用意すれば良いですか? Q4. キャピラリーシーケンサが自施設に無い場合でも実験はできますか? Q5. FFPE 検体でも実験はできますか? 2. 実験のセットアップ Q1. 実験において重要なポイントはありますか?
Microsoft PowerPoint - DNA1.ppt [互換モード]
![Microsoft PowerPoint - DNA1.ppt [互換モード]](/thumbs/94/118241118.jpg "Microsoft PowerPoint - DNA1.ppt [互換モード]") 生物物理化学 タンパク質をコードする遺伝子 (135~) 本 PPT 資料の作成には福岡大学機能生物研究室のホームページを参考にした http://133.100.212.50/~bc1/biochem/index2.htm 1 DA( デオキシリボ核酸 ) の化学的特徴 シャルガフ則とDAのX 線回折像をもとに,DAの構造が予測された (Watson & Crick 1953 年 ) 2 Watson
生物物理化学 タンパク質をコードする遺伝子 (135~) 本 PPT 資料の作成には福岡大学機能生物研究室のホームページを参考にした http://133.100.212.50/~bc1/biochem/index2.htm 1 DA( デオキシリボ核酸 ) の化学的特徴 シャルガフ則とDAのX 線回折像をもとに,DAの構造が予測された (Watson & Crick 1953 年 ) 2 Watson
Gen とるくん™(酵母用)High Recovery
High Recovery") 研究用 Gen とるくん ( 酵母用 ) High Recovery 説明書 v201510da Gen とるくん ( 酵母用 )High Recovery は 細胞壁分解酵素による酵母菌体処理と塩析による DNA 精製の組み合わせにより 効率良く酵母ゲノム DNA を抽出 精製するためのキットです 本キットを用いた酵母ゲノム DNA 調製操作は 遠心による酵母菌体の回収 GenTLE Yeast
研究用 Gen とるくん ( 酵母用 ) High Recovery 説明書 v201510da Gen とるくん ( 酵母用 )High Recovery は 細胞壁分解酵素による酵母菌体処理と塩析による DNA 精製の組み合わせにより 効率良く酵母ゲノム DNA を抽出 精製するためのキットです 本キットを用いた酵母ゲノム DNA 調製操作は 遠心による酵母菌体の回収 GenTLE Yeast
テイカ製薬株式会社 社内資料
 テイカ製薬株式会社社内資料 アレルギー性結膜炎治療剤トラニラスト点眼液.5% TS TRANILAST Ophthalmic Solution.5% TS 生物学的同等性に関する資料 発売元 : 興和株式会社 製造販売元 : テイカ製薬株式会社 9 年 月作成 TSTR5BE9 ラット及びモルモットアレルギー性結膜炎モデルにおける生物学的同等性試験 Ⅰ. 試験の目的トラニラスト点眼液.5% TS および標準製剤の生物学的同等性をラット受動感作アレルギー性結膜炎モデル及びモルモット能動感作アレルギー性結膜炎モデルを用い薬力学的に検討した
テイカ製薬株式会社社内資料 アレルギー性結膜炎治療剤トラニラスト点眼液.5% TS TRANILAST Ophthalmic Solution.5% TS 生物学的同等性に関する資料 発売元 : 興和株式会社 製造販売元 : テイカ製薬株式会社 9 年 月作成 TSTR5BE9 ラット及びモルモットアレルギー性結膜炎モデルにおける生物学的同等性試験 Ⅰ. 試験の目的トラニラスト点眼液.5% TS および標準製剤の生物学的同等性をラット受動感作アレルギー性結膜炎モデル及びモルモット能動感作アレルギー性結膜炎モデルを用い薬力学的に検討した
本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因
 HbA1c 測定系について ~ 原理と特徴 ~ 一般社団法人日本臨床検査薬協会 技術運営委員会副委員長 安部正義 本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因 HPLC 法 HPLC 法原理 高速液体クロマトグラフィー 混合物の分析法の一つ 固体または液体の固定相 ( 吸着剤 ) 中で 液体または気体の移動相 ( 展開剤 ) に試料を加えて移動させ
HbA1c 測定系について ~ 原理と特徴 ~ 一般社団法人日本臨床検査薬協会 技術運営委員会副委員長 安部正義 本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因 HPLC 法 HPLC 法原理 高速液体クロマトグラフィー 混合物の分析法の一つ 固体または液体の固定相 ( 吸着剤 ) 中で 液体または気体の移動相 ( 展開剤 ) に試料を加えて移動させ
Hi-level 生物 II( 国公立二次私大対応 ) DNA 1.DNA の構造, 半保存的複製 1.DNA の構造, 半保存的複製 1.DNA の構造 ア.DNA の二重らせんモデル ( ワトソンとクリック,1953 年 ) 塩基 A: アデニン T: チミン G: グアニン C: シトシン U
 DNA 1.DNA の構造, 半保存的複製 1.DNA の構造, 半保存的複製 1.DNA の構造 ア.DNA の二重らせんモデル ( ワトソンとクリック,1953 年 ) 塩基 A: アデニン T: チミン G: グアニン C: シトシン U") 1.DNA の構造, 半保存的複製 1.DNA の構造 ア.DNA の二重らせんモデル ( ワトソンとクリック,1953 年 ) 塩基 A: アデニン T: チミン G: グアニン C: シトシン U: ウラシル (RNA に含まれている塩基 DNA にはない ) イ. シャルガフの規則 二本鎖の DNA に含まれる A,T,G,C の割合は,A=T,G=C となる 2.DNA の半保存的複製 ア.
1.DNA の構造, 半保存的複製 1.DNA の構造 ア.DNA の二重らせんモデル ( ワトソンとクリック,1953 年 ) 塩基 A: アデニン T: チミン G: グアニン C: シトシン U: ウラシル (RNA に含まれている塩基 DNA にはない ) イ. シャルガフの規則 二本鎖の DNA に含まれる A,T,G,C の割合は,A=T,G=C となる 2.DNA の半保存的複製 ア.
タンパク質は 電気泳動後にタンパク質を可視化するための最も簡単で迅速な方法です バイオ ラッドは感度 定量性 質量分析計対応に合わせた可視 蛍光剤を取り扱っています CBB クマシーブリリアントブルーは を行ったゲル中のタンパク質をするための最も一般的な方法です バイオ ラッドでは 用途に合わせて2
 ゲル剤 バイオ ラッドでは ゲル中のタンパク質を検出する剤を数多く取り扱っています 感度 操作性 再現性 定量性 電気泳動後のアプリケーションへの影響など サンプルと目的に応じて最適な剤は異なります 剤を選択する際に 下記の剤セレクションガイドをご参照ください 剤セレクションガイド 製品名 用途 プロセス数 所要時間目安 最高感度 事前調製 特徴 CBB Bio-Safe CBB G-250 ステイン
ゲル剤 バイオ ラッドでは ゲル中のタンパク質を検出する剤を数多く取り扱っています 感度 操作性 再現性 定量性 電気泳動後のアプリケーションへの影響など サンプルと目的に応じて最適な剤は異なります 剤を選択する際に 下記の剤セレクションガイドをご参照ください 剤セレクションガイド 製品名 用途 プロセス数 所要時間目安 最高感度 事前調製 特徴 CBB Bio-Safe CBB G-250 ステイン
論文題目 腸管分化に関わるmiRNAの探索とその発現制御解析
 論文題目 腸管分化に関わる microrna の探索とその発現制御解析 氏名日野公洋 1. 序論 microrna(mirna) とは細胞内在性の 21 塩基程度の機能性 RNA のことであり 部分的相補的な塩基認識を介して標的 RNA の翻訳抑制や不安定化を引き起こすことが知られている mirna は細胞分化や増殖 ガン化やアポトーシスなどに関与していることが報告されており これら以外にも様々な細胞諸現象に関与していると考えられている
論文題目 腸管分化に関わる microrna の探索とその発現制御解析 氏名日野公洋 1. 序論 microrna(mirna) とは細胞内在性の 21 塩基程度の機能性 RNA のことであり 部分的相補的な塩基認識を介して標的 RNA の翻訳抑制や不安定化を引き起こすことが知られている mirna は細胞分化や増殖 ガン化やアポトーシスなどに関与していることが報告されており これら以外にも様々な細胞諸現象に関与していると考えられている
日本標準商品分類番号 カリジノゲナーゼの血管新生抑制作用 カリジノゲナーゼは強力な血管拡張物質であるキニンを遊離することにより 高血圧や末梢循環障害の治療に広く用いられてきた 最近では 糖尿病モデルラットにおいて増加する眼内液中 VEGF 濃度を低下させることにより 血管透過性を抑制す
 日本標準商品分類番号 872491 カリジノゲナーゼの血管新生抑制作用 カリジノゲナーゼは強力な血管拡張物質であるキニンを遊離することにより 高血圧や末梢循環障害の治療に広く用いられてきた 最近では 糖尿病モデルラットにおいて増加する眼内液中 VEGF 濃度を低下させることにより 血管透過性を抑制することが示されたが 血管新生に対するカリジノゲナーゼの影響を評価した報告はない そこで今回 網膜血管新生に対するカリジノゲナーゼの役割を同定するため
日本標準商品分類番号 872491 カリジノゲナーゼの血管新生抑制作用 カリジノゲナーゼは強力な血管拡張物質であるキニンを遊離することにより 高血圧や末梢循環障害の治療に広く用いられてきた 最近では 糖尿病モデルラットにおいて増加する眼内液中 VEGF 濃度を低下させることにより 血管透過性を抑制することが示されたが 血管新生に対するカリジノゲナーゼの影響を評価した報告はない そこで今回 網膜血管新生に対するカリジノゲナーゼの役割を同定するため
大学院委員会について
 博士学位請求論文 審査報告書 審査委員 ( 主査 ) 農学部専任准教授村上周一朗 ( 副査 ) 農学部専任教授中島春紫 ( 副査 ) 農学部専任准教授前田理久 2014 年 1 月 17 日 1 論文提出者氏名高橋結 2 論文題名 ( 邦文題 ) ハイイロジェントルキツネザルの糞由来 Aspergillus niger E-1 株の有する xylanase に関する研究 ( 欧文訳 )Study on
博士学位請求論文 審査報告書 審査委員 ( 主査 ) 農学部専任准教授村上周一朗 ( 副査 ) 農学部専任教授中島春紫 ( 副査 ) 農学部専任准教授前田理久 2014 年 1 月 17 日 1 論文提出者氏名高橋結 2 論文題名 ( 邦文題 ) ハイイロジェントルキツネザルの糞由来 Aspergillus niger E-1 株の有する xylanase に関する研究 ( 欧文訳 )Study on
遺伝子検査の基礎知識
 遺伝子検査の準備と注意事項 PCR はわずか 1 分子の鋳型 DNA でも検出可能であるため 反応液調製時に鋳型となりうる核酸等が混入しないように細心の注意が必要です この文書では 正確な遺伝子検査を行うために必要な実験器具類やコンタミネーション防止のための注意事項について解説します - 目次 - 1 実験環境の整備 2 必要な実験器具と装置 1 実験環境の整備 コンタミネーションの原因 PCR は非常に高感度な検出法であるため
遺伝子検査の準備と注意事項 PCR はわずか 1 分子の鋳型 DNA でも検出可能であるため 反応液調製時に鋳型となりうる核酸等が混入しないように細心の注意が必要です この文書では 正確な遺伝子検査を行うために必要な実験器具類やコンタミネーション防止のための注意事項について解説します - 目次 - 1 実験環境の整備 2 必要な実験器具と装置 1 実験環境の整備 コンタミネーションの原因 PCR は非常に高感度な検出法であるため
PrimeScript® II 1st strand cDNA Synthesis Kit
 研究用 PrimeScript II 1st strand cdna Synthesis Kit 説明書 v201208da 目次 I. 概要... 3 II. キットの内容... 4 III. 保存... 4 IV. 1st-strand cdna 合成反応... 5 V. RT-PCR を行う場合... 6 VI. RNA サンプルの調製について... 6 VII. 関連製品... 7 VIII.
研究用 PrimeScript II 1st strand cdna Synthesis Kit 説明書 v201208da 目次 I. 概要... 3 II. キットの内容... 4 III. 保存... 4 IV. 1st-strand cdna 合成反応... 5 V. RT-PCR を行う場合... 6 VI. RNA サンプルの調製について... 6 VII. 関連製品... 7 VIII.
Western BLoT Rapid Detect
 研究用 Western BLoT Rapid Detect 説明書 v201212 Western BLoT Rapid Detect は 標識二次抗体の代わりに独自の IgG Detector(HRP labeled) を利用して一次抗体を検出するウェスタンブロッティング専用の検出試薬キットです 本製品を利用することで 標識二次抗体を用いて検出する従来法ではできなかった迅速検出 高感度検出 シグナルの増強
研究用 Western BLoT Rapid Detect 説明書 v201212 Western BLoT Rapid Detect は 標識二次抗体の代わりに独自の IgG Detector(HRP labeled) を利用して一次抗体を検出するウェスタンブロッティング専用の検出試薬キットです 本製品を利用することで 標識二次抗体を用いて検出する従来法ではできなかった迅速検出 高感度検出 シグナルの増強
PowerPoint プレゼンテーション
 透析液ライン管理のエキスパートを目指して 第 20 回兵庫県臨床工学技士会定期学習会 臨床現場における透析液ラインの洗浄消毒法選択とその効果 課題 過酢酸を含む消毒剤の選択 田岡正宏 1 目的 1) 消毒の効果 薬剤の種類の 2 剤化 2) 消毒時間の短縮化 1 日の使用薬剤の 1 剤化 使用薬剤の選択 透析液ライン管理に用いられる洗浄剤の種類と特徴 過酢酸を含む消毒剤の組成 過酢酸濃度順 過酢酸
透析液ライン管理のエキスパートを目指して 第 20 回兵庫県臨床工学技士会定期学習会 臨床現場における透析液ラインの洗浄消毒法選択とその効果 課題 過酢酸を含む消毒剤の選択 田岡正宏 1 目的 1) 消毒の効果 薬剤の種類の 2 剤化 2) 消毒時間の短縮化 1 日の使用薬剤の 1 剤化 使用薬剤の選択 透析液ライン管理に用いられる洗浄剤の種類と特徴 過酢酸を含む消毒剤の組成 過酢酸濃度順 過酢酸
NGS_KAPA RNA HyperPrep Kit
 シークエンシングワークフロー ライブラリー調製 サンプル 調製 末端修復 エンドポイントライブラリー増幅 A-TAILING アダプター ライゲーション サイズセレクション & サイズ確認 または リアルタイムライブラリー増幅 ライブラリー 定量 クラスター 増幅 KAPA RNA HyperPrep Kit illumina社用ライブラリー調製キット KAPA RNA HyperPrep Kit
シークエンシングワークフロー ライブラリー調製 サンプル 調製 末端修復 エンドポイントライブラリー増幅 A-TAILING アダプター ライゲーション サイズセレクション & サイズ確認 または リアルタイムライブラリー増幅 ライブラリー 定量 クラスター 増幅 KAPA RNA HyperPrep Kit illumina社用ライブラリー調製キット KAPA RNA HyperPrep Kit
<4D F736F F D B82C982C282A282C482512E646F63>
 サンプル条件および固定化分子の選択 Biacoreの実験ではセンサーチップに固定化する分子をリガンド それに対して結合を測定する分子をアナライトと呼びます いずれの分子をリガンドとし アナライトとするかは 実験系を構築する上で重要です 以下にサンプルに適したリガンド アナライトの設計方法やサンプルの必要条件などをご紹介します アナライト リガンド センサーチップ (1) タンパク質リガンドとしてもアナライトとしても用いることができます
サンプル条件および固定化分子の選択 Biacoreの実験ではセンサーチップに固定化する分子をリガンド それに対して結合を測定する分子をアナライトと呼びます いずれの分子をリガンドとし アナライトとするかは 実験系を構築する上で重要です 以下にサンプルに適したリガンド アナライトの設計方法やサンプルの必要条件などをご紹介します アナライト リガンド センサーチップ (1) タンパク質リガンドとしてもアナライトとしても用いることができます
 ,...~,.'~ 表 2.6.2.2-26 試験管内 PAE 菌株薬剤 MIC (µg/ml) PAE (h) 1 MIC 4 MIC STFX 0.025 0.92 2.35 S. aureus FDA 209-P LVFX 0.20 0.68 2.68 CPFX 0.20 1.05 1.59 SPFX 0.10 0.35 1.07 STFX 0.025 2.33 1.14 E. coli KL-16
,...~,.'~ 表 2.6.2.2-26 試験管内 PAE 菌株薬剤 MIC (µg/ml) PAE (h) 1 MIC 4 MIC STFX 0.025 0.92 2.35 S. aureus FDA 209-P LVFX 0.20 0.68 2.68 CPFX 0.20 1.05 1.59 SPFX 0.10 0.35 1.07 STFX 0.025 2.33 1.14 E. coli KL-16
様式 C-19 科学研究費助成事業 ( 科学研究費補助金 ) 研究成果報告書 平成 25 年 5 月 16 日現在 機関番号 : 研究種目 : 基盤研究 (C) 研究期間 : 2010 ~ 2012 課題番号 : 研究課題名 ( 和文 ) ルーメン内における水素生成微生
 研究成果報告書 平成 25 年 5 月 16 日現在 機関番号 : 研究種目 : 基盤研究 (C) 研究期間 : 2010 ~ 2012 課題番号 : 研究課題名 ( 和文 ) ルーメン内における水素生成微生") Kaken Contents Title / 科研費報告書 ( 構成タイトル ) ルーメン内における水素生成微生物および 水素利用微生物の分子生態学的解析 Molecular ecological analysis o hydrogen-utilizing microorganis 松井, 宏樹 MATSUI, HIROKI 三重大学, 2013. 2010 年度 ~2012 年度科学研究費補助金
Kaken Contents Title / 科研費報告書 ( 構成タイトル ) ルーメン内における水素生成微生物および 水素利用微生物の分子生態学的解析 Molecular ecological analysis o hydrogen-utilizing microorganis 松井, 宏樹 MATSUI, HIROKI 三重大学, 2013. 2010 年度 ~2012 年度科学研究費補助金
2009年度業績発表会(南陽)
") 高速イオンクロマトグラフィーによる ボイラ水中のイオン成分分析 のご紹介 東ソー株式会社 バイオサイエンス事業部 JASIS 217 新技術説明会 (217.9.8) rev.1 1. ボイラ水分析について ボイラ水の水質管理 ボイラ : 高圧蒸気の発生装置であり 工場, ビル, 病院など幅広い産業分野でユーティリティ源として利用されている 安全かつ効率的な運転には 日常の水質管理, ブロー管理が必須
高速イオンクロマトグラフィーによる ボイラ水中のイオン成分分析 のご紹介 東ソー株式会社 バイオサイエンス事業部 JASIS 217 新技術説明会 (217.9.8) rev.1 1. ボイラ水分析について ボイラ水の水質管理 ボイラ : 高圧蒸気の発生装置であり 工場, ビル, 病院など幅広い産業分野でユーティリティ源として利用されている 安全かつ効率的な運転には 日常の水質管理, ブロー管理が必須
2017 年 2 月 27 日 Myco Finder バリデーションデータ 日水製薬株式会社 研究部
 2017 年 2 月 27 日 Myco Finder バリデーションデータ 日水製薬株式会社 研究部 1. 概要 マイコプラズマ遺伝子検出キット (Myco Finder) の性能を評価するために 下記ふたつの情報を参 考にバリデーションデータを取得した 1. 第十七改正日本薬局方 ( 平成 28 年 3 月 7 日厚生労働省告示 64 号 ) 参考情報 バイオテクノロジー応用医薬品 / 生物起源由来医薬品の製造に用いる細胞基材に対するマイコ
2017 年 2 月 27 日 Myco Finder バリデーションデータ 日水製薬株式会社 研究部 1. 概要 マイコプラズマ遺伝子検出キット (Myco Finder) の性能を評価するために 下記ふたつの情報を参 考にバリデーションデータを取得した 1. 第十七改正日本薬局方 ( 平成 28 年 3 月 7 日厚生労働省告示 64 号 ) 参考情報 バイオテクノロジー応用医薬品 / 生物起源由来医薬品の製造に用いる細胞基材に対するマイコ
スライド 1
 第 56 回日本透析医学会 2011.06.19 血液ガス分析装置による透析液重炭酸イオン濃度 と総二酸化炭素濃度 ~ その正確性について ~ 五仁会元町 HD クリニック臨床検査部 同臨床工学部 * 同内科 ** 清水康 田中和弘 小松祐子 森上辰哉 * 田中和馬 * 阪口剛至 * 大槻英展 * 吉本秀之 * 田渕篤嗣 * 申曽洙 ** 目的 各社血液ガス分析装置を用いて重炭酸イオン ( ) を
第 56 回日本透析医学会 2011.06.19 血液ガス分析装置による透析液重炭酸イオン濃度 と総二酸化炭素濃度 ~ その正確性について ~ 五仁会元町 HD クリニック臨床検査部 同臨床工学部 * 同内科 ** 清水康 田中和弘 小松祐子 森上辰哉 * 田中和馬 * 阪口剛至 * 大槻英展 * 吉本秀之 * 田渕篤嗣 * 申曽洙 ** 目的 各社血液ガス分析装置を用いて重炭酸イオン ( ) を
井上先生 報告書 清水
 派遣研究室 : ボン大学 Life and Medical Sciences(LIMES) Institute, Prof.Michael Hoch 研究室 研究 交流概要近年 TAL effector nuclease (TALEN) や CRISPR/Cas9 システムが効率的で簡便なゲノム編集技術として注目されている 派遣者は TALEN を用いてゼブラフィッシュに遺伝子変異を導入する実験を行った
派遣研究室 : ボン大学 Life and Medical Sciences(LIMES) Institute, Prof.Michael Hoch 研究室 研究 交流概要近年 TAL effector nuclease (TALEN) や CRISPR/Cas9 システムが効率的で簡便なゲノム編集技術として注目されている 派遣者は TALEN を用いてゼブラフィッシュに遺伝子変異を導入する実験を行った
Cytotoxicity LDH Assay Kit-WST
 ytotoxicity L ssay Kit-WST はじめに 本説明書は ytotoxicity L ssay Kit-WST を用いた抗体依存性細胞傷害測定用 (ntibody-dependent cellmediated cytotoxicity: ) です 本製品のキット内容や Working Solution の調製方法に関して 製品添付の取扱い説明書も合わせてご覧ください 正確な測定のために
ytotoxicity L ssay Kit-WST はじめに 本説明書は ytotoxicity L ssay Kit-WST を用いた抗体依存性細胞傷害測定用 (ntibody-dependent cellmediated cytotoxicity: ) です 本製品のキット内容や Working Solution の調製方法に関して 製品添付の取扱い説明書も合わせてご覧ください 正確な測定のために
Taro-04-1(雌雄判別)
") 高嶋康晴 Yasuharu TAKASHIMA 要 約 多くの PCR 法では 電気泳動により分離された特定塩基長の PCR 産物や制限酵素で断片化された DNA 断片の塩基長のパターンを目視で検出し生物種や品種等の判定を行う 電気泳動にかかる一連の操作を手作業で行うことによる分析者への負担の軽減及び分析処理の効率化を図るために マグロ属魚類及びサケ科魚類の魚種判別法について アガロースゲル電気泳動と全自動電気泳動装置による電気泳動の比較検討を行った
高嶋康晴 Yasuharu TAKASHIMA 要 約 多くの PCR 法では 電気泳動により分離された特定塩基長の PCR 産物や制限酵素で断片化された DNA 断片の塩基長のパターンを目視で検出し生物種や品種等の判定を行う 電気泳動にかかる一連の操作を手作業で行うことによる分析者への負担の軽減及び分析処理の効率化を図るために マグロ属魚類及びサケ科魚類の魚種判別法について アガロースゲル電気泳動と全自動電気泳動装置による電気泳動の比較検討を行った
ノーウオ―クウイルスのPCR法
 厚生労働科学研究新型インフルエンザ等新興 再興感染症研究事業 現在 国内で分離 同定できないウイルス性出血熱等の診断等の対応方法に関する研究 班より SFTS ウイルス検査マニュアル 平成 25 年 3 月 13 日 RT-PCR 法により SFTS ウイルス核蛋白質 (NP) 遺伝子を特異的に検出 同定する検査法を解説する 本法は 1 本の反応チューブ内で逆転写反応 遺伝子増幅を連続的に行い SFTS
厚生労働科学研究新型インフルエンザ等新興 再興感染症研究事業 現在 国内で分離 同定できないウイルス性出血熱等の診断等の対応方法に関する研究 班より SFTS ウイルス検査マニュアル 平成 25 年 3 月 13 日 RT-PCR 法により SFTS ウイルス核蛋白質 (NP) 遺伝子を特異的に検出 同定する検査法を解説する 本法は 1 本の反応チューブ内で逆転写反応 遺伝子増幅を連続的に行い SFTS
(Microsoft Word - \202\205\202\2232-1HP.doc)
") イーズ NO.002(15 年 8 月発行 ) 日本薬局方 微生物限度試験法微生物限度試験法の実際 監修 : 国立衛生試験所三瀬勝利 前号では微生物限度試験法について 全体的な解説を行ったが 本号では生菌数試験 大腸菌 サルモネラ 緑膿菌 黄色ブドウ球菌の試験項目ごとに それぞれ実際の試験の流れを図解してみた 試験を行う際に役立てて頂ければ幸いである また無菌試験法についても補冊として添付したので参照されたい
イーズ NO.002(15 年 8 月発行 ) 日本薬局方 微生物限度試験法微生物限度試験法の実際 監修 : 国立衛生試験所三瀬勝利 前号では微生物限度試験法について 全体的な解説を行ったが 本号では生菌数試験 大腸菌 サルモネラ 緑膿菌 黄色ブドウ球菌の試験項目ごとに それぞれ実際の試験の流れを図解してみた 試験を行う際に役立てて頂ければ幸いである また無菌試験法についても補冊として添付したので参照されたい
DNA Fragmentation Kit
 研究用 DNA Fragmentation Kit 説明書 v201712da 本製品は 超音波破砕装置などの特殊な装置を使用せず 酵素処理によってゲノム DNA 等の長鎖 DNA をランダムに断片化し さらに平滑化するためのキットです 平滑化した断片はそのまま平滑末端ベクターに組み込むことができます また 平滑化を必要としない場合は 断片化までで反応を止めることもできます メチル化 DNA 濃縮や高速シーケンス解析のための前処理にも
研究用 DNA Fragmentation Kit 説明書 v201712da 本製品は 超音波破砕装置などの特殊な装置を使用せず 酵素処理によってゲノム DNA 等の長鎖 DNA をランダムに断片化し さらに平滑化するためのキットです 平滑化した断片はそのまま平滑末端ベクターに組み込むことができます また 平滑化を必要としない場合は 断片化までで反応を止めることもできます メチル化 DNA 濃縮や高速シーケンス解析のための前処理にも
PrimeSTAR® GXL DNA Polymerase
 研究用 PrimeSTAR GXL DNA Polymerase 説明書 v201308da PrimeSTAR GXL DNA Polymerase は PrimeSTAR HS DNA Polymerase を改良し さらに独自の伸長因子を組み合わせることにより PCR パフォーマンスを飛躍的に向上させた 画期的な High Fidelity PCR 酵素です 非常に高い正確性を維持しながら これまでの
研究用 PrimeSTAR GXL DNA Polymerase 説明書 v201308da PrimeSTAR GXL DNA Polymerase は PrimeSTAR HS DNA Polymerase を改良し さらに独自の伸長因子を組み合わせることにより PCR パフォーマンスを飛躍的に向上させた 画期的な High Fidelity PCR 酵素です 非常に高い正確性を維持しながら これまでの
解析法
 1.Ct 値の算出方法 Ct 値の算出方法には 閾値と増幅曲線の交点を Ct 値とする方法 (Crossing Point 法 ) の他に 増幅曲線の 2 次導関数を求めてそれが最大となる点を Ct 値とする方法がある (2nd Derivative Maximum 法 ) 前者では 閾値を指数関数的増幅域の任意の位置に設定して解析するが その位置により Ct 値が変化するので実験間の誤差が大きくなりやすい
1.Ct 値の算出方法 Ct 値の算出方法には 閾値と増幅曲線の交点を Ct 値とする方法 (Crossing Point 法 ) の他に 増幅曲線の 2 次導関数を求めてそれが最大となる点を Ct 値とする方法がある (2nd Derivative Maximum 法 ) 前者では 閾値を指数関数的増幅域の任意の位置に設定して解析するが その位置により Ct 値が変化するので実験間の誤差が大きくなりやすい
はじめに
 LAMP 法を用いたウェルシュ菌エンテロトキシン遺伝子検出の検討 佐藤秀美 Development of detection of Clostridium perfringens enterotoxin gene using the LAMP assey Hidemi Sato はじめにウェルシュ菌による食中毒は, 大量に調理された食品を原因とすることが多く, しばしば大規模になるため, 原因物質の迅速な検出と判断が求められている.
LAMP 法を用いたウェルシュ菌エンテロトキシン遺伝子検出の検討 佐藤秀美 Development of detection of Clostridium perfringens enterotoxin gene using the LAMP assey Hidemi Sato はじめにウェルシュ菌による食中毒は, 大量に調理された食品を原因とすることが多く, しばしば大規模になるため, 原因物質の迅速な検出と判断が求められている.
はじめてのリアルタイムPCR
 はじめてのリアルタイム PCR 1. はじめにリアルタイム PCR 法は PCR 増幅産物の増加をリアルタイムでモニタリングし 解析する技術です エンドポイントで PCR 増幅産物を確認する従来の PCR 法に比べて 1DNA や RNA の正確な定量ができること 2 電気泳動が不要なので迅速かつ簡便に解析できコンタミネーションの危険性も小さいことなど多くの利点があります 今や 遺伝子発現解析や SNP
はじめてのリアルタイム PCR 1. はじめにリアルタイム PCR 法は PCR 増幅産物の増加をリアルタイムでモニタリングし 解析する技術です エンドポイントで PCR 増幅産物を確認する従来の PCR 法に比べて 1DNA や RNA の正確な定量ができること 2 電気泳動が不要なので迅速かつ簡便に解析できコンタミネーションの危険性も小さいことなど多くの利点があります 今や 遺伝子発現解析や SNP
PowerPoint プレゼンテーション
 酵素 : タンパク質の触媒 タンパク質 Protein 酵素 Enzyme 触媒 Catalyst 触媒 Cataylst: 特定の化学反応の反応速度を速める物質 自身は反応の前後で変化しない 酵素 Enzyme: タンパク質の触媒 触媒作用を持つタンパク質 第 3 回 : タンパク質はアミノ酸からなるポリペプチドである 第 4 回 : タンパク質は様々な立体構造を持つ 第 5 回 : タンパク質の立体構造と酵素活性の関係
酵素 : タンパク質の触媒 タンパク質 Protein 酵素 Enzyme 触媒 Catalyst 触媒 Cataylst: 特定の化学反応の反応速度を速める物質 自身は反応の前後で変化しない 酵素 Enzyme: タンパク質の触媒 触媒作用を持つタンパク質 第 3 回 : タンパク質はアミノ酸からなるポリペプチドである 第 4 回 : タンパク質は様々な立体構造を持つ 第 5 回 : タンパク質の立体構造と酵素活性の関係
Word Pro - matome_7_酸と塩基.lwp
 酸と 酸と 酸 acid 亜硫酸 pka =.6 pka =.9 酸 acid ( : 酸, すっぱいもの a : 酸の, すっぱい ) 酸性 p( ) 以下 酸っぱい味 ( 酸味 ) を持つ リトマス ( ) BTB( ) 金属と反応して ( ) を発生 ( 例 )Z l Zl リン酸 P pka =.5 pka =. pka =.8 P P P P P P P 酸性のもと 水素イオン 塩化水素
酸と 酸と 酸 acid 亜硫酸 pka =.6 pka =.9 酸 acid ( : 酸, すっぱいもの a : 酸の, すっぱい ) 酸性 p( ) 以下 酸っぱい味 ( 酸味 ) を持つ リトマス ( ) BTB( ) 金属と反応して ( ) を発生 ( 例 )Z l Zl リン酸 P pka =.5 pka =. pka =.8 P P P P P P P 酸性のもと 水素イオン 塩化水素
Western BLoT Immuno Booster
 研究用 Western BLoT Immuno Booster 説明書 v201211 Western BLoT Immuno Booster は 抗体の反応性を増強させる成分を含む溶液で 抗体の希釈液に用いるだけで 抗原抗体反応を促進します 本製品は ウェスタンブロット ELISA 等の各種イムノアッセイに対応しており 各アッセイにおいて数倍から数十倍の検出感度向上が期待できます 西洋ワサビペルオキシダーゼ
研究用 Western BLoT Immuno Booster 説明書 v201211 Western BLoT Immuno Booster は 抗体の反応性を増強させる成分を含む溶液で 抗体の希釈液に用いるだけで 抗原抗体反応を促進します 本製品は ウェスタンブロット ELISA 等の各種イムノアッセイに対応しており 各アッセイにおいて数倍から数十倍の検出感度向上が期待できます 西洋ワサビペルオキシダーゼ
LA PCR™i n vitro Cloning Kit
 研究用 LA PCR in vitro Cloning Kit 説明書 v201201da 本キットは Cassette および Cassette Primer を使用することにより cdna やゲノム上の未知領域を特異的に増幅させる従来の PCR in vitro Cloning Kit に 長鎖 DNA を効率良く増幅する LA PCR Technology を応用したシステムです これにより
研究用 LA PCR in vitro Cloning Kit 説明書 v201201da 本キットは Cassette および Cassette Primer を使用することにより cdna やゲノム上の未知領域を特異的に増幅させる従来の PCR in vitro Cloning Kit に 長鎖 DNA を効率良く増幅する LA PCR Technology を応用したシステムです これにより