学位申請論文 岡山大学大学院医歯薬学総合研究科 病態制御科学 病態機構学講座歯周病態学分野 田口裕子 ( 平成 26 年 12 月 8 日受付 ) 1
|
|
|
- しのぶ おおかわち
- 5 years ago
- Views:
Transcription
1 学位申請論文 岡山大学大学院医歯薬学総合研究科 病態制御科学 病態機構学講座歯周病態学分野 田口裕子 ( 平成 26 年 12 月 8 日受付 ) 1
2 Porphyromonas gingivalis における菌体外のジンジパインの 酵素活性に関わる新規遺伝子について 田口裕子 Yuko Taguchi 2
3 緒言 歯周病は細菌の混合感染により引き起こされる慢性炎症性疾患であり, 歯肉の炎症と歯槽骨の破壊を引き起こす 1) グラム陰性偏性嫌気性細菌 Porphyromonas gingivalis は歯周病の主たる原因菌として考えられており, 莢膜, 線毛,lipopolysaccharide(LPS), そして強力なプロテアーゼを主な病原因子として有する 2) P. gingivalis は複数のプロテアーゼを持つが, なかでもジンジパインは強力なプロテアーゼであり, 直接または間接的に歯周組織を破壊する 3, 4) ジンジパインには,2 種類のアルギニンジンジパイン (RgpA, RgpB) とリジンジンジパイン (Kgp) の3 種類がある 5, 6) Kgp および RgpA は, シグナルペプチド, プロペプチド,Lys プロテアーゼドメインまたは Arg プロテアーゼドメイン, 付着ドメイン, および C 末端ドメイン (C-terminal domain:ctd) からなる 一方,RgpB は, シグナルペプチド, プロペプチド,Arg プロテアーゼドメイン, および CTD からなる Kgp と Rgp は C 末端に CTD を持つことから,CTD 蛋白質に位置づけられる CTD とは, 菌体表層に局在する一部の蛋白質の C 末端に存在する約 80アミノ酸からなるドメインで,CTD 蛋白質の分泌や, 菌体表層への局在に関与する P. gingivalis では, 約 34 個の CTD 蛋白質が存在する 7) CTD 蛋白質は近年その存在が明らかとなった Type IX(Por) 分泌装置 (T9SS) から分泌されることが明らかになっている 8) CTD 蛋白質は, 細胞質で合成された後, まず内膜にある Sec 分泌装置を通過してペリプラズムへと移行する その後,T9SS を通過して菌体表層でプロセッシングされ, 菌体外へ分泌されるか, もしくは, 菌体表層で外膜へ固定される T9SS は複数の蛋白質によって構築されている装置であり,PorK,PorL,PorM,PorN,PorP,PorQ,PorT, PorU,PorV(PG27,LptO),PorW, そして Sov から構成される 9, 10) CTD は CTD 蛋白質が T9SS を通過した後に切断されるドメインであり,T9SS を通過するために必要なペプチド配列と考えられている そして,T9SS を通過した CTD 蛋白質には陰性に荷電 3
4 したリポ多糖である anionic lipopolysaccharide(a-lps) が修飾される 11) ところで,Escherichia coli をはじめとする複数のグラム陰性細菌において, 外膜蛋白質は, 内膜から外膜にまで輸送される際に,SurA (peptidyl-prolyl isomerase),skp {OmpH (Outer membrane protein H)}, そして DegP{high temperature requirement A(HtrA)} をはじめとするシャペロンの助けを必要とすることが報告されている 12) P. gingivalis においては HtrA についての報告はあるが,SurA と Skp(OmpH) に関する報告はこれまでなされていない P. gingivalis のゲノム配列上には htra に加え,surA と omph のホモログも存在する P. gingivalis ATCC33277 株のゲノム上に存在する omph のホモログである PGN_0300の遺伝子産物である PGN_0300 蛋白質が,P. gingivalis の CTD 蛋白質の成熟と修飾に関与している可能性を考え, 研究を行った 4
5 材料および方法 1. 使用菌株, 使用プラスミド, および培養条件本研究に使用した菌株とプラスミドは, 表 1と表 2にそれぞれ記した プラスミドの作製には E. coli DH5α 株を使用した E. coli の培養にはアンピシリン100 µg/ml 含有 Luria-Bertain(LB) 培地 ( ナカライテスク, 京都 ) を使用し,37 C の好気条件下で培養した P. gingivalis の培養には, 変法 BHI 培地 {Brain Heart Infusion(BHI)(Becton,Dickinson and Company(BD),Franklin Lakes,NJ,USA),Yeast Extract(BD), ヘミン ( 和光純薬工業, 大阪 ), メナジオン ( 和光純薬工業 ), および L-システイン塩酸塩 1 水和物 (Sigma) を含む } 5), 変法 Tryptic soy(ts) 寒天培地 {Tryptic soy agar(bd),bhi, ヘミン, メナジオン, および L-システイン塩酸塩 1 水和物を含む } 5), および血液寒天培地 { 緬羊脱繊維血液 ( 日本バイオテスト研究所, 東京 ), ヘミン, メナジオン, および L-システイン塩酸塩 1 水和物を含む } を用いた なお,PGN_0300を欠損させた株(PGN_0300 欠損株 ) の培養には, エリスロマイシン (Em)10 µg/ml を,PGN_0300 欠損株に PGN_0300 を再度保有させた株 (PGN_0300 相補株 ) の培養には Em 10 µg/ml およびテトラサイクリン (Tc)0.7 µg/ml を培地に添加して使用した 培養には, 嫌気ボックス (Whitley Workstation DG250,Microbiology International,Frederick,MD,USA) を用い,37 C の嫌気条件下 (10% CO 2,10% H 2,80% N 2 ) で培養した 2. 遺伝子操作制限酵素はタカラバイオ ( 滋賀 ) で購入した 核酸の泳動用緩衝液には1 Tris-acetate-ethylenediaminetetraacetic acid(edta)( TAE,1mM EDTA を含む40 mm トリス酢酸緩衝液 ;ph 8.0) を使用し,1% または2% アガロースゲルを用いた 分離し 5
6 た核酸断片は臭化エチジウム (0.5 mg/ml) で染色し, ゲル撮影装置 Dopphin-View( ク ラボウ, 東京 ) を使用して, 紫外線 ( 波長 :320nm) 照射下にて撮影した 3.PGN_0300 欠損株と PGN_0300 相補株の作製 PGN_0300 欠損株の作製は,PGN_0300 遺伝子座に Em 耐性遺伝子カセットを挿入することによって作製した P. gingivalis ATCC33277 株のゲノム DNA を鋳型とし,PGN_0300の上流および下流領域各 1,000 bp をプライマーセット PGN_0300U-F と PGN_0300U-R, および PGN_0300D-F と PGN_0300D-R を使用し,PrimeSTAR GXL DNA polymerase( タカラバイオ ) を用いて polymerase chain reaction (PCR) 法により増幅した アガロースゲル電気泳動法によって目的の大きさの DNA 断片が増幅されていることを確認後,PCR 産物を QIA quick PCR Purification Kit(Qiagen,Hilden,Germany) を用いて精製した 上流および下流領域の PCR 産物をそれぞれ EcoRI と BamHI あるいは XbaI と BamHI を用いて37 C で3 時間消化し,Mini Elute Reaction Cleanup Kit(Qiagen) で精製した 精製した2 種類の DNA 断片と,EcoRI と XbaI で制限酵素処理した puc19を,dna Ligation Kit <Mighty Mix>( タカラバイオ ) を用いて16 C で1 時間反応させることにより結合させ, その後 E. coli DH5α 株に形質転換した puc19に2つの断片が正しく挿入されたプラスミドを選択し, その BamHI サイトに Em 耐性遺伝子カセットを挿入した Em 耐性遺伝子カセットは,Em 耐性遺伝子カセットが挿入されたプラスミド pkd355 13) を鋳型として, プライマーセット Em-F と Em-R を用いて PCR 法によって増幅した 得られた PCR 産物を,QIA quick PCR Purification Kit を用いて精製した後に BamHI で消化し,PGN_0300 領域が挿入された上記のプラスミドに組み込んだ E. coli DH5α 株に形質転換後,Em 耐性遺伝子カセットが正方向に挿入されたプラスミド puc-pgn_0300を精製し,puvii で線形化した後, P. gingivalis ATCC33277 株に電気穿孔法 (1.25 kv/mm) によって導入した Em を含む血 6
7 液寒天培地で集落を形成した菌について, ゲノム上の PGN_0300 遺伝子と Em 耐性遺伝子カセットが正しく置換されたことを PGN_0300 遺伝子座と Em 耐性遺伝子カセットの配列特異的プライマーを用いた PCR 法で確認した PGN_0300 相補株は,PGN_1045 遺伝子座に PGN_0300 遺伝子を挿入することによって作製した PGN_1045はβ-ガラクトシダーゼをコードしており,P. gingivalis はラクトースを利用しないため, この遺伝子を欠損させても生存に影響しないと考えられるため選択した PGN_0300 相補株作製のために,P. gingivalis ATCC33277 株のゲノム DNA を鋳型とし, PGN_1045の上流および下流領域の各 1,000 bp をプライマーセット PGN_1045U-F と PGN_1045U-R, および PGN_1045D-F と PGN_1045D-R を使用し,PrimeSTAR GXL DNA polymerase を用いて PCR 法によって増幅した 精製した上流および下流領域の PCR 産物をそれぞれ XhoI と HindIII, BamHI と NotI により37 C で3 時間消化したのち,Mini Elute Reaction Cleanup Kit で精製した 上流領域を pbluescript II SK(-)(Agilent Technologies, Santa Clara, CA, USA) の XhoI-HindIII サイトに挿入して, プラスミド pb-1045a を得た そして,pB-1045A を BamHI と NotI で消化し, 制限酵素処理した PGN_1045 下流領域 DNA 断片を挿入してプラスミド pb-1045ab を得た 次に,pB-1045AB を BamHI で消化し,BamHI と BglII を用いてプラスミド pkd375 14) を切断して得た Tc 耐性遺伝子カセットを挿入して pk-1045tet を得た pk-1045tet に fima プロモーターを挿入してプラスミド pcpg を作製した このために,P. gingivalis ATCC33277 株のゲノム DNA を鋳型とし,fimA プロモーター領域をプライマーセット FimA-PRO-F と FimA-PRO-R を用いて PCR 法で増幅し,StuI と BamHI で消化した後,p1045TET の SmaI-BamHI 切断サイトに挿入した 最後に P. gingivalis ATCC33277 株のゲノム DNA を鋳型とし, プライマーセット PGN_0300ORF-F と PGN_0300ORF-R で増幅した PGN_0300の Open Reading Frame を BamHI で消化した後,pCPG の fima プロモーター直下の BamHI サイトに挿入 7
8 して, プラスミド pcpg-pgn_0300を構築した BssHII を用いて pcpg-pgn_0300を線形化したのち,pgn_0300 欠損株へ電気穿孔法にて導入した Em および Tc を含む血液寒天培地上に集落を形成した菌を用いて, ゲノム上の PGN_1045 遺伝子座と, 導入した pcpg-pgn_0300の DNA 断片との間で正しく相同組換えが行われていることを PGN_1045 遺伝子座の配列特異的プライマーを用いた PCR 法で確認した 4. 逆転写ポリメラーゼ連鎖反応 (RT-PCR) 全 RNA の抽出は TRIzol(Invitrogen, Carlsbad, CA, USA) を用いて行った 精製した RNA サンプルは,DNaseI( タカラバイオ ) を用いて37 C で1 時間処理した後, ゲノム DNA の混入がないことを, プライマーセット PGN_0299-F と PGN_0299-R を用いた PCR 法で増幅産物が生じないことで確認した 2 µg の RNA を SuperscriptIII First Strand Synthesis Supermix(Invitrogen) とランダムヘキサマープライマー (Invitrogen) を用いて cdna に逆転写した 得られた cdna を鋳型として,PrimeSTAR GXL DNA Polymerase, プライマーセット PGN_0299-F と PGN_0299-R,PGN_0300-F と PGN_0300-R, および PGN_0301-F と PGN_0301-R を使用した PCR 法を行うことで,PGN_0299, PGN_0300, および PGN_0301の各遺伝子の発現を確認した 5. プロテアーゼ活性の測定 P. gingivalis は,BHI 培地で37 C, 嫌気下で一晩培養した 菌体と培養上清は4 C, 10,000 g で20 分間遠心して分離した 集菌した菌体はリン酸緩衝液 (Invitrogen) で懸濁した Kgp と Rgp の活性測定は Sato らの方法に準じて行い 9), それぞれに特異的な蛍光基質である Benzyloxcarbonyl-L-phenylalanyl-L-arginine 4-methyl-coumaryl-7-amide(Z-Phe-Arg- MCA ) と Benzyloxycarbonyl-L-histidyl-L-glutamyl-L-lysine 4-Methyl-coumaryl-7-amide 8
9 (Z-His-Glu- Lys-MCA)( ペプチド研究所, 大阪 ) を用いた 培養上清および菌体に5 mm システインを含有する20 mm リン酸緩衝液 (ph 7.0) と蛍光基質を加え,40 C で10 分間反応させ,460 nm の蛍光波長 ( 励起波長 380 nm) で測定した Kgp と Rgp の活性は, 1 分間に1 nmol のアミノメチルクマリンを遊離する活性を1ユニットと定義した 6. 細胞分画 P. gingivalis の菌体成分は,Sato らの方法 15) に準じて分画した すなわち,200 ml の P. gingivalis 培養物を4 C で,10,000 g,10 分間遠心して, 菌体と培養上清に分けた 菌体を100 ml のリン酸緩衝液 (0.1mM N α -p-tosyl-l-lysine chloromethyl ketone および0.1 mm leupeptin を含有する ) で懸濁し, フレンチプレスを用いて100 MPa の加圧下で2 回処理して菌体を破砕した 破砕されなかった菌体は,4 C で2,400 g,10 分間遠心して取り除いた その上清を4 C で,100,000 g,1 時間遠心した後, その上清画分を細胞質とペリプラズム画分として回収した 沈澱物は,1% Triton X-100と20 mm MgCl 2 を含有するリン酸緩衝液で懸濁し,30 C で20 分間静置した その後,4 C で100,000 g,1 時間遠心した後, 上清画分を内膜画分として回収し, 沈澱物を外膜画分として回収した ベジクル非含有の培養上清を得るためには,140 ml の P. gingivalis 培養物を4 C で, 6,000 g,10 分間遠心した 次に得られた上清をさらに4 C で100,000 g,1 時間遠心分離して, この上清をベジクル非含有培養上清とした ベジクル非含有培養上清中の蛋白質は, 最終濃度 20% になるようにトリクロロ酢酸を加え, 氷上で1 時間静置することによって沈殿させ, 沈澱物をアセトンで2 回洗浄した 洗浄した沈殿物を乾燥させた後,0.1 mm N α -p-tosyl-l-lysine chloromethyl ketone(tlck) と0.1 mm ロイペプチン含有の滅菌蒸留水に溶解した 7. 抗体 9
10 PGN_0300 蛋白質に対する抗体の作製には,PGN_0300 蛋白質の部分ペプチドを使用した すなわち,N 末端にシステイン残基を付与した PGN_0300の膜貫通ドメイン以外の領域中の93 番目のイソロイシンから106 番目のリジンまでに相当するペプチド CIVKKEQQASELKRK を合成し, ヘモシアニン (KLH) を共有結合させた ( オペロンバイオテクノロジーへ委託, 東京 ) モルモットへの免疫と採血はイブバイオサイエンス ( 和歌山 ) へ委託した 抗 PGN_0300ペプチドモルモット抗血清は,1/500 希釈で使用した 抗 Kgp ウサギポリクローナル抗体 16) と抗 Rgp マウスポリクローナル抗体 16) は,1/5,000 希釈で使用した A-LPS を認識するマウスモノクローナル抗体 1B5 17) は,Curtis 博士より恵与していただいた 8. ウエスタンブロット法ドデシル硫酸ナトリウム (SDS) ポリアクリルアミドゲル電気泳動 (PAGE) は Laemmli 18) らの方法に従い行った 泳動に用いる試料は,2 mm オルトバナジウム酸塩および1 mm フッ化ナトリウムを含有する SDS サンプルバッファーに溶解した 還元状態で95 C,10 分間の加熱により変性させた後, ポリアクリルアミドを含むゲルと泳動用緩衝液 (25 mm Tris-HCl,200 mm グリシン,35 mm SDS) を用いて泳動して蛋白質を分離した 分離した蛋白質を湿式転写装置 (Mini Trans-Blot Cell; Bio-Rad) を用い, 転写用緩衝液 (25 mm Tris HCl,192 mm グリシン,20% メタノール ) 中で polyvinylidene difluoride(pvdf) 膜へ転写した (100 V 定電圧下 ) 転写後の PVDF 膜を5% スキムミルク (BD) 含有 Tris-Tween 緩衝食塩水 (TBST 緩衝液 ;20 mm Tris-HCl,pH 7.6,150 mm NaCl, および0.05% Tween20) に1 時間浸してブロッキング操作を行った その後,1 次抗体を5% スキムミルク含有 TBST 緩衝液で前述の濃度に希釈した溶液中に PVDF 膜を浸漬し,4 C で12 時間反応させた 反応後, 1% スキムミルク含有 TBST 緩衝液で1 時間洗浄し, 次いで2 次抗体を5% スキムミルク含 10
11 有 TBST 緩衝液で希釈した溶液中に PVDF 膜を浸漬し, 常温で1 時間反応させた 2 次抗体として horseradish peroxidase(hrp) で標識した抗ウサギ IgG 抗体 (Dako, Glostrup, Denmark), 抗マウス IgG 抗体 (Dako), あるいは抗モルモット IgG 抗体 (Millipore Co., Temecula,CA,USA) を用いた 1% スキムミルク含有 TBST 緩衝液で1 時間洗浄後,ECL Western Blotting Detection Reagents(GE Healthcare, Uppsala, Sweden) および ChemiDoc MP (Bio Lad) を用いて検出を行った 9. 統計処理 各実験系における統計解析には, 対応のない群間の Student s t test を用いた なお,p 値が 0.05 以下をもって有意差ありと判定した 11
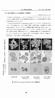
12 結果 1.PGN_0300の菌体内局在 P. gingivalis ATCC33277 株のゲノム上には,National Center for Biotechnology Information(NCBI) データーベースによると,2つの omph ホモログである PGN_0300 と PGN_0301が存在する PGN_0300と PGN_0301のアミノ酸の相同性は24.9% であった ( 図 1) PGN_0300と PGN_0301のアミノ酸配列を TMHMM Server v. 2.0 software ( で解析することによって, 両蛋白質が膜蛋白質である可能性を検討した その結果,PGN_0300 蛋白質は,N 末端 ( Arg 7 から Leu 29 ) に膜貫通ドメインを1つ有することが予測され, 膜蛋白質である可能性が高いことが示唆された 一方,PGN_0301 蛋白質には膜貫通ドメインの存在は予測されなかった PGN_0300 蛋白質の局在を確認するために,P. gingivalis ATCC33277 株の菌体成分を分画し, ウエスタンブロット法で抗 PGN_0300ペプチド抗体と反応させた その結果, 外膜画分に約 17 kda のバンドが強く検出された ( 図 2) 以上のことから PGN_0300 蛋白質も外膜蛋白質であることが推測された 2.PGN_0300 欠損株の作製 P. gingivalis ATCC33277 株のゲノム上の PGN_0300 遺伝子を Em 耐性遺伝子カセットに置換した PGN_0300 欠損株は, 血液寒天培地上で白色コロニーを形成した また, PGN_0300 相補株は, 親株と同様に黒色コロニーを形成した ( 図 3) PGN_0300 欠損株における PGN_0300 遺伝子の発現の消失および,PGN_0300 遺伝子を欠失したことによる前後の PGN_0299 遺伝子と PGN_0301 遺伝子の発現への影響を,RT-PCR 法で調べた 親株,PGN_0300 欠損株, および PGN_0300 相補株から抽出した全 RNA から調整した cdna を鋳型とし,PGN_0300 遺伝子特異的プライマーセットを用いて PCR を行ったところ, 12
13 親株および PGN_0300 相補株の cdna では約 360 bp の増幅産物が得られた ( 図 4) これに対して PGN_0300 欠損株の cdna を用いた場合には増幅産物は全くなかった このことから,PGN_0300 欠損株では PGN_0300 遺伝子は発現されていないことが確認された PGN_0299 遺伝子あるいは PGN_0301 遺伝子それぞれに特異的なプライマーを用いた場合には, 親株,PGN_0300 欠損株, および PGN_0300 相補株のいずれの cdna においても約 450 bp あるいは約 350 bp の増幅産物がそれぞれ検出された ( 図 4) このことから PGN_0300 欠損株において, PGN_0300 遺伝子の破壊は PGN_0299 遺伝子および PGN_0301 遺伝子の発現に影響を及ぼしていないことが確認された 以上のことから, PGN_0300 欠損株における白色コロニーの形成は,PGN_0300 遺伝子の欠損が原因であることが示された 3.PGN_0300 欠損株におけるジンジパインの活性 PGN_0300 欠損株におけるジンジパインの活性を検討した PGN_0300 欠損株の菌体と培養上清中における Kgp と Rgp の活性を調べたところ, 菌体と培養上清中のいずれにおいても Kgp と Rgp の活性はともに顕著に低下していた 一方,PGN_0300 相補株の Kgp と Rgp の活性は, 菌体および培養上清中のいずれにおいても, ともに部分的に回復していた ( 図 5) 4. 菌体および培養上清における Kgp と Rgp 存在様式の解析 PGN_0300 欠損株では, 菌体および培養上清中における Kgp と Rgp の活性が低下していたことから, 菌体および培養上清中の Kgp と Rgp の存在様式を, 抗 Kgp 抗体と抗 Rgp 抗体を用いたウエスタンブロット法で解析した 菌体の全蛋白質を用いたところ,Kgp および Rgp の蛋白質量は親株に対し,PGN_0300 欠損株では顕著に少なかった ( 図 6) PGN_0300 欠損株では特に成熟型の Kgp と推測される約 50 kda の蛋白質の量が顕著に低 13
14 下しており, 逆にプロ型と推測される約 190 kda の蛋白質の比率が増加していた また, Rgp では糖鎖修飾されたと考えられる約 kda のスメアな蛋白質の量および, 成熟型の Rgp と推測される43 kda の蛋白質の量が PGN_0300 欠損株では顕著に低下していた 培養上清中の蛋白質に対して抗 Kgp 抗体を反応させたところ, 親株の培養上清では成熟型の Kgp と推測される約 50 kda の蛋白質が検出された 一方,PGN_0300 欠損株の培養上清では成熟型 Kgp は検出されず, 未成熟型の約 140 kda の蛋白質の比率が増加していた また, 抗 Rgp 抗体との反応においても, 親株の培養上清では成熟型 Rgp と推定される約 43 kda の蛋白質が検出されたのに対して, 欠損株の培養上清では未成熟型 Rgp と推定される75 kda 以上の蛋白質が顕著に検出されたが, 成熟型 Rgp はわずかに検出されたのみであった PGN_0300 相補株における反応はいずれの場合においても親株に対する反応に類似していた 5. 菌体内における Kgp と Rgp の局在親株と PGN_0300 欠損株における Kgp と Rgp の局在を比較した 親株と PGN_0300 欠損株の菌体を, 菌体全抽出物, 細胞質 ペリプラズム画分, 内膜 外膜画分, 内膜画分, 外膜画分にそれぞれ分画し, 抗 Kgp 抗体と抗 Rgp 抗体を用いたウエスタンブロット法で解析した ( 図 7) 親株では, 成熟型 Kgp と推定される約 50 kda の蛋白質が外膜画分に検出された しかし PGN_0300 欠損株では, 外膜画分を含めたいずれの画分においても成熟型 Kgp は検出されず, 未成熟型の Kgp と推定される約 190 kda の蛋白質が菌体全抽出物とともに細胞質 ペリプラズム画分に検出された また, 親株では, 成熟型 RgpA と推定される約 43 kda の蛋白質とともに, 糖鎖修飾された成熟型 RgpB と推定される蛋白質が60-80 kda にスメアなバンドとして外膜画分に検出された これに対し,PGN_0300 欠損株では, 成熟型 RgpB は外膜画分を含めたい 14
15 ずれの画分においても検出されず, 未成熟型 RgpA と推定される約 120 kda の蛋白質と, 未成熟型 RgpB と推定される約 70 kda の蛋白質が菌体全抽出物とともに細胞質 ペリプ ラズム画分に検出された 6.A-LPS の検出親株,PGN_0300 欠損株, および PGN_0300 相補株における A-LPS の存在を,A-LPS に反応するモノクローナル抗体 1B5を用いたウエスタンブロット法で検討した 図 8に示すように, いずれの株においても1B5と反応するスメアなバンドが検出された しかし, 親株と PGN_0300 相補株ではスメアなバンドが約 200 kda の高分子にまでおよんでいたのに対し,PGN_0300 欠損株では約 70 kda までにとどまっていた 15
16 考察 本研究では,P. gingivalis ATCC33277 株を親株として作製した PGN_0300 欠損株が, 血 液寒天培地上で白色コロニーを形成したことから,PGN_0300 蛋白質はジンジパインで ある Kgp と Rgp の活性に関与していることが強く示唆された P. gingivalis は血液寒天 培地上で黒色コロニーを形成する これは, ヘモグロビンが分解された際に遊離される ヘムが菌体表面上で µ-oxo heme dimer として蓄積することによる 19) ヘモグロビンの分 解にはジンジパインが関与し,kgp の遺伝子変異株では血液寒天培地上でコロニーの黒 色化が減弱し, 白色コロニーとなる 20) kgp に変異が無いにも関わらず白色コロニーを形成する変異株が複数報告されている 15, 21-34) 15) 23) これらの欠損株はその表現型から,porT 型と porr 型に分けられる port 型の欠損株では, 菌体と上清のいずれにおいてもジンジパイン活性がなく,Kgp と Rgp を含む CTD 蛋白質が菌体外へ分泌されず, ペリプラズムに蓄積する 一方,porR 型の欠損株では,CTD 蛋白質は分泌されるが, 菌体のジンジパイン活性は消失する しかし 上清中のジンジパイン活性は維持される porr 型の欠損株では A-LPS が合成されないため 24-26, 28-30, 34) 菌体表層における CTD 蛋白質への A-LPS 修飾が行われず, その結果,CTD 蛋白質は菌体表層へ固定されない port 型と porr 型以外に,LptO 31) と PorU 32) の変異が報告されている LptO の欠損株では,A-LPS は合成されるが, ペリプラズムに蓄積し,CTD 蛋白質には結合しない 31) 一方で PorU は CTD 蛋白質から CTD を切断するペプチダーゼであり 32),PorU 欠損株では CTD が切断されないため, 菌体表層へ分泌された CTD 蛋白質は A-LPS は結合しない その結果,CTD 蛋白質は外膜には固定されず, 菌体外へ放出される PGN_0300 欠損株はその外膜画分に抗 Kgp 抗体あるいは抗 Rgp 抗体と反応する蛋白質は全く検出されなかった そして, 未成熟型と考えられる Kgp と Rgp が細胞質あるいはペリプラズムに蓄積していた ( 図 7) また, 培養上清 16
17 中にもプロ型と考えられる未成熟の Kgp と Rgp が検出された 1B5 抗体を用いたウエスタンブロット法では, PGN_0300 欠損株の全菌体において,A-LPS のスメアなバンドが検出されたが, 低分子側にシフトしており A-LPS は CTD 蛋白質非結合型であることが示された 以上の PGN_0300 欠損株の表現型は,PorU 欠損株の表現型に類似していることが示唆された PGN_0300 欠損株において CTD 蛋白質における CTD の切断が行われない可能性に加え,A-LPS の輸送が正常には行われていない可能性も残されている 今後, この点を明らかにする必要がある ところで,CTD 蛋白質の多くは外膜蛋白質である 複数のグラム陰性菌において, SurA, Skp(OmpH), あるいは DegP(HtrA) をはじめとする分子シャペロンが外膜蛋白質に対して作用することが報告されている 35, 36) これらは, 外膜蛋白質が誤った構造を取ることや凝集することを防ぎ, 正しい構造を取るようにフォールディングすることを助けている P. gingivalis ATCC33277 株のゲノム上にも sura,degp(htra), そして omph ホモログが存在する しかし,P. gingivalis の SurA や Skp(OmpH) に関する報告はない PGN_0300と PGN_0301は OmpH ホモログである これまでの報告では,Skp (OmpH) は菌種毎にその局在が異なっている E. coli では Skp は膜貫通ドメインを有しておらず, ペリプラズムに局在し, 外膜蛋白質がペリプラズムを通過する際にシャペロンとして作用し, それらのβバレル構造の形成を助ける機能がある 35) Flavobacterium psychrophilum 37), や Yersinia pseudotuberculosis 38) では OmpH として外膜に局在している 細菌における OmpH の機能は不明であるが, 外膜にあることに関連し, 抗原性が高いことが示されている 37, 38) 本研究において TMHMM Server v. 2.0 software による解析の結果,PGN_0300 蛋白質は膜貫通ドメインを1つ有する外膜蛋白質であることが予測され, 抗 PGN_0300ペプチド抗体を用いたウエスタンブロット法の結果からも外膜に局在することが支持された 本研究を総括すると,PGN_0300は外膜蛋白質であり,T9SS から分泌された Kgp, Rgp 17
18 のフォールディングに関与し, それらの形状を正常に保つことに関与している可能性があげられる CTD は CTD 蛋白質が T9SS によって分泌されるとき, また,CTD 切断後に CTD 蛋白質に A-LPS が修飾されるときにシグナルとして機能することが示唆されている そして,CTD が機能するためには, その一次構造よりも立体構造が重要であることが報告されている 39) この立体構造を維持するために PGN_0300 蛋白質が機能している可能性がある あるいは CTD を切断する PorU に働き, その機能を正常に保つために作用している可能性がある PorU は CTD 蛋白質のひとつであるが, 他の CTD 蛋白質とは異なり外膜に運ばれたのちにも CTD が切断されない特殊なタイプの CTD 蛋白質である 32) PGN_0300が PorU に特異的に働いている可能性がある 今後は,PGN_0300 と PGN_0301がシャペロンとして機能をしている可能性を含め,PGN_0300がどのようなメカニズムで, ジンジパインの分泌, プロセッシングに関与しているかを明らかにする必要がある 18
19 結論 P. gingivalis の OmpH 様蛋白質である PGN_0300 蛋白質は, ジンジパインの分泌, プロ セッシング, および糖鎖修飾に関与することが示唆された 謝辞 稿を終えるにあたり, 懇篤なる御指導, 御高閲を賜りました岡山大学大学院医歯薬学総合研究科口腔微生物学分野, 大原直也教授, そして主任教授であります岡山大学大学院医歯薬学総合研究科歯周病態学分野, 高柴正悟教授に謹んで感謝の意を表します また, 実験試料を恵与いただきましたとともに貴重な御助言を頂きました長崎大学大学院医歯薬学総合研究科口腔病原微生物学分野, 中山浩次教授, 内藤真理子准教授, 佐藤啓子助教, 雪竹英治技術職員に感謝申し上げます 1B5 抗体を恵与いただきましたロンドン大学クイーンメアリー校 Michael A. Curtis 教授に感謝申し上げます 最後に本研究を行うにあたり, 貴重な御助言を頂きました岡山大学大学院医歯薬学総合研究科口腔微生物分野および歯周病態学分野の諸先生方に厚く御礼申し上げます 19
20 文献 1)R.C. Page, S. Offenbacher, H.E. Schroeder, G.J. Seymour, K.S. Kornman. Advances in the pathogenesis of periodontitis: summary of developments, clinical implications and future directions, Periodontol (1997) )S.C. Holt, J.L. Ebersole. Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia: the "red complex", a prototype polybacterial pathogenic consortium in periodontitis, Periodontol (2005) ) J. Potempa, A. Banbula, J. Travis. Role of bacterial proteinases in matrix destruction and modulation of host responses, Periodontol (2000) )E. Andrian, Y. Mostefaoui, M. Rouabhia, D. Grenier. Regulation of matrix metalloproteinases and tissue inhibitors of matrix metalloproteinases by Porphyromonas gingivalis in an engineered human oral mucosa model, J Cell Physiol 211 (2007) )K. Nakayama, T. Kadowaki, K. Okamoto, K. Yamamoto. Construction and characterization of arginine-specific cysteine proteinase (Arg-gingipain)-deficient mutants of Porphyromonas gingivalis. Evidence for significant contribution of Arg-gingipain to virulence, J Biol Chem 270 (1995) )J. Potempa, R. Pike, J. Travis. The multiple forms of trypsin-like activity present in various strains of Porphyromonas gingivalis are due to the presence of either Arg-gingipain or Lys-gingipain, Infect Immun 63 (1995) )C.A. Seers, N. Slakeski, P.D. Veith, T. Nikolof, Y.Y. Chen, S.G. Dashper, E.C. Reynolds. The RgpB C-terminal domain has a role in attachment of RgpB to the outer membrane and belongs to a novel C-terminal-domain family found in Porphyromonas gingivalis, J Bacteriol 188 (2006) )K. Sato, H. Yukitake, Y. Narita, M. Shoji, M. Naito, K. Nakayama. Identification of Porphyromonas gingivalis proteins secreted by the Por secretion system, FEMS Microbiol Lett 338 (2013) )K. Sato, M. Naito, H. Yukitake, H. Hirakawa, M. Shoji, M.J. McBride, R.G. Rhodes, K. Nakayama. A protein secretion system linked to bacteroidete gliding motility and pathogenesis, Proc Natl Acad Sci U S A 107 (2010) )M. Shoji, K. Sato, H. Yukitake, Y. Kondo, Y. Narita, T. Kadowaki, M. Naito, K. Nakayama. Por secretion system-dependent secretion and glycosylation of Porphyromonas gingivalis hemin-binding protein 35, PLoS One 6 (2011) e
21 11)N. Slakeski, C.A. Seers, K. Ng, C. Moore, S.M. Cleal, P.D. Veith, A.W. Lo, E.C. Reynolds. C-terminal domain residues important for secretion and attachment of RgpB in Porphyromonas gingivalis, J Bacteriol 193 (2011) )J.G. Sklar, T. Wu, D. Kahne, T.J. Silhavy. Defining the roles of the periplasmic chaperones SurA, Skp, and DegP in Escherichia coli, Genes Dev 21 (2007) ) J. Ueshima, M. Shoji, D.B. Ratnayake, K. Abe, S. Yoshida, K. Yamamoto, Nakayama, K. Purification, gene cloning, gene expression, and mutants of Dps from the obligate anaerobe Porphyromonas gingivalis, Infect Immun 71 (2003) ) Y. Shi, D.B. Ratnayake, K. Okamoto, N. Abe, K. Yamamoto, K. Nakayama. Genetic analyses of proteolysis, hemoglobin binding, and hemagglutination of Porphyromonas gingivalis. Construction of mutants with a combination of rgpa, rgpb, kgp, and haga, J Biol Chem 274 (1999) )K. Sato, E. Sakai, P.D. Veith, M. Shoji, Y. Kikuchi, H. Yukitake, N. Ohara, M. Naito, K. Okamoto, E.C. Reynolds, K. Nakayama. Identification of a new membrane-associated protein that influences transport/maturation of gingipains and adhesins of Porphyromonas gingivalis, J Biol Chem 280 (2005) ) R. Takii, T. Kadowaki, A. Baba, T. Tsukuba, K. Yamamoto. A functional virulence complex composed of gingipains, adhesins, and lipopolysaccharide shows high affinity to host cells and matrix proteins and escapes recognition by host immune systems, Infect Immun 73 (2005) )M.A. Curtis, A. Thickett, J.M. Slaney, M. Rangarajan, J. Aduse-Opoku, P. Shepherd, N. Paramonov, E.F. Hounsell. Variable carbohydrate modifications to the catalytic chains of the RgpA and RgpB proteases of Porphyromonas gingivalis W50, Infect Immun 67 (1999) )U.K. Laemmli. Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature 227 (1970) )J.W. Smalley, J. Silver, P.D. Marsh, A.J. Birss. The periodomtopathogen Porphyromonas gingivalis binds iron protoporphyrin IX in the µ-oxo dimeric form: an oxidive buffer and possible pathogenic mechanism, Biochem J 331 (1998) )K. Okamoto, K. Nakayama, T. Kadowaki, N. Abe, D.B. Ratnayake, K. Yamamoto. Involvement of a lysine-specific cysteine proteinase in hemoglobin adsorption and heme accumulation by Porphyromonas gingivalis, J Biol Chem 273 (1998) )C.I. Hoover, F. Yoshimura. Transposon-induced pigment-deficient mutants of Porphyromonas gingivalis, FEMS Microbiol Lett 124 (1994)
22 22)T. Chen, H. Dong, R. Yong, M.J. Duncan. Pleiotropic pigmentation mutants of Porphyromonas gingivalis, Microb Pathog 28 (2000) )M. Shoji, D.B. Ratnayake, Y. Shi, T. Kadowaki, K. Yamamoto, F. Yoshimura, A. Akamine, M.A. Curtis, K. Nakayama. Construction and characterization of a nonpigmented mutant of Porphyromonas gingivalis: cell surface polysaccharide as an anchorage for gingipains, Microbiology 148 (2002) )E. Vanterpool, F. Roy, H.M. Fletcher. The vime gene downstream of vima is independently expressed and is involved in modulating proteolytic activity in Porphyromonas gingivalis W83, Infect Immun 72 (2004) )E. Vanterpool, F. Roy, H.M. Fletcher. Inactivation of vimf, a putative glycosyltransferase gene downstream of vime, alters glycosylation and activation of the gingipains in Porphyromonas gingivalis W83, Infect Immun 73 (2005) )E. Vanterpool, F. Roy, L. Sandberg, H.M. Fletcher. Altered gingipain maturation in vimaand vime-defective isogenic mutants of Porphyromonas gingivalis, Infect Immun 73 (2005) )E. Vanterpool, F. Roy, W. Zhan, S.M. Sheets, L. Sangberg, H.M. Fletcher. VimA is part of the maturation pathway for the major gingipains of Porphyromonas gingivalis W83, Microbiology 152 (2006) )N.A. Paramonov, J. Aduse-Opoku, A. Hashim, M. Rangarajan, M.A. Curtis. Structural analysis of the core region of O-lipopolysaccharide of Porphyromonas gingivalis from mutants defective in O-antigen ligase and O-antigen polymerase, J Bacteriol 191 (2009) )K. Sato, N. Kido, Y. Murakami, C.I. Hoover, K. Nakayama, F. Yoshimura. Lipopolysaccharide biosynthesis-related genes are required for colony pigmentation of Porphyromonas gingivalis, Microbiology 155 (2009) )M. Yamaguchi, K. Sato, H. Yukitake, Y. Noiri, S. Ebisu, K. Nakayama. A Porphyromonas gingivalis mutant defective in a putative glycosyltransferase exhibits defective biosynthesis of the polysaccharide portions of lipopolysaccharide, decreased gingipain activities, strong autoaggregation, and increased biofilm formation, Infect Immun 78 (2010) )Y.Y. Chen, B. Peng, Q. Yang, M.D. Glew, P.D. Veith, K.J. Cross, K.N. Goldie, D. Chen, N. O'Brien-Simpson, S.G. Dashper, E.C. Reynolds. The outer membrane protein LptO is essential for the O-deacylation of LPS and the co-ordinated secretion and attachment of A-LPS and CTD proteins in Porphyromonas gingivalis, Mol Microbiol 79 (2011) )M.D. Glew, P.D. Veith, B. Peng, Y.Y. Chen, D.G. Gorasia, Q. Yang, N. Slakeski, D. Chen, C. 22
23 Moore, S. Crawford, E.C. Reynolds. PG0026 is the C-terminal signal peptidase of a novel secretion system of Porphyromonas gingivalis, J Biol Chem 287 (2012) )M. Shoji, H. Yukitake, K. Sato, Y. Shibata, M. Naito, J. Aduse-Opoku, Y. Abiko, M.A. Curtis, K. Nakayama. Identification of an O-antigen chain length regulator, WzzP, in Porphyromonas gingivalis, Microbiologyopen 2 (2013) )M. Shoji, K. Sato, H. Yukitake, M. Naito, K. Nakayama. Involvement of the Wbp pathway in the biosynthesis of Porphyromonas gingivalis lipopolysaccharide with anionic polysaccharide, Sci Rep 4 (2014) )C. Goemans, K. Denoncin, J.F. Collet. Folding mechanisms of periplasmic proteins, Biochim Biophys Acta 1843 (2014) )B. Lipinska, M. Zylicz, C. Georgopoulos. The HtrA (DegP) protein, essential for Escherichia coli survival at high temperatures, is an endopeptidase, J Bacteriol 172 (1990) )F. Dumetz, E. Duchaud, S.E. LaPatra, C. Le Marrec, S. Claverol, M.C. Urdaci, M. Le Henaff. A protective immune response is generated in rainbow trout by an OmpH-like surface antigen (P18) of Flavobacterium psychrophilum, Appl Environ Microbiol 72 (2006) )R. Vuorio, L. Hirvas, R.B. Raybourne, D.T. Yu, M. Vaara. The nucleotide and deduced amino acid sequence of the cationic 19 kda outer membrane protein OmpH of Yersinia pseudotuberculosis, Biochim Biophys Acta 1129 (1991) )X.Y. Zhou, J.L. Gao, N. Hunter, J. Potempa, K.A. Nguyen. Sequence-independent processing site of the C-terminal domain (CTD) influences maturation of the RgpB protease from Porphyromonas gingivalis, Mol Microbiol 89 (2013)
24 表題脚注 岡山大学大学院医歯薬学総合研究科病態制御科学専攻病態機構学講座 歯周病態学分野 ( 指導高柴正悟教授 ) 岡山大学大学院医歯薬学総合研究科社会環境生命科学専攻国際環境科学講座 口腔微生物学分野 ( 委託大原直也教授 ) 本論文の一部は, 以下の学会において発表した 第 56 回歯科基礎医学会学術大会総会 (2014 年 9 月, 福岡 ) 24
25 図の説明 図 1.PGN_0300と PGN_0301のアミノ酸配列の比較 P. gingivalis ATCC33277 株のゲノムに存在する omph ホモログ PGN_0300と PGN_0301 のアミノ酸配列を比較した 同一のアミノ酸, 保存的アミノ酸置換, 半保存的アミノ酸置換をそれぞれアスタリスク, コロン, およびドットで示す ダッシュは配列を最大限に一致させたときに生じたギャップを示す 図 2.PGN_0300 蛋白質の局在 P. gingivalis ATCC33277 株の菌体抽出物を分画し, 抗 PGN_0300 蛋白質ペプチド抗体を用いたウエスタンブロット法で PGN_0300 蛋白質の局在を調べた M, 分子量マーカー ;1, 全菌体抽出物 ;2, 細胞質 ペリプラズム画分 ;3, 内膜 外膜画分 ;4, 内膜画分 ;5, 外膜画分 ;CBB 染色, クーマシーブリリアントブルー染色 図 3. コロニー性状の比較血液寒天培地上における P. gingivalis ATCC33277 株,PGN_0300 欠損株, および PGN_0300 相補株のコロニー性状を比較した 1,P. gingivalis ATCC 株 ;2,PGN_0300 欠損株 ;3,PGN_0300 相補株 図 4. PGN_0299, PGN_0300, および PGN_0301の各遺伝子の発現 P. gingivalis ATCC33277 株,PGN_0300 欠損株, および PGN_0300 相補株の cdna を鋳型とし,PGN_0299, PGN_0300, PGN_0301の発現を確認した M,100 bp ラダー ;1,P. gingivalis ATCC33277 株 ;2,PGN_0300 欠損株 ;3,PGN_0300 相補株 図 5. ジンジパインの活性 全菌体あるいは培養上清と,Kgp あるいは Rgp に対する蛍光基質を 40 C で 10 分間反 25
26 応させたのちに蛍光波長を測定した 縦軸は P. gingivalis ATCC33277 株における活性を 100% とした場合の,PGN_0300 欠損株と PGN_0300 相補株における Kgp あるいは Rgp の活性を示す なお, エラーバーは標準偏差を示す 1,P. gingivalis ATCC33277 株 ;2,PGN_0300 欠損株 ;3,PGN_0300 相補株 図 6. 菌体と培養上清中の Kgp と Rgp 形状の解析抗 Kgp 抗体と抗 Rgp 抗体を用いたウエスタンブロット法により, 菌体および培養上清中の Kgp と Rgp を検出した M, 分子量マーカー ;1,P. gingivalis ATCC33277 株 ;2,PGN_0300 欠損株 ;3,PGN_0300 相補株 図 7.Kgp と Rgp の菌体内局在 P. gingivalis ATCC33277 株および PGN_0300 欠損株の菌体抽出物を分画し, 抗 Kgp 抗体と抗 Rgp 抗体を用いたウエスタンブロット法を行い,Kgp と Rgp の局在を明らかにした M, 分子量マーカー ;1, 全菌体抽出物 ;2, 細胞質 ペリプラズム画分 ;3, 内膜 外膜画分 ;4, 内膜画分 ;5, 外膜画分 ;CBB 染色, クーマシーブリリアントブルー染色 図 8. 菌体中の A-LPS の形状の解析 A-LPS を認識するモノクローナル抗体 1B5 抗体を菌体抽出物と反応させた 1,P. gingivalis ATCC33277 株 ;2,PGN_0300 欠損株 ;3,PGN_0300 相補株 26
プロトコール集 ( 研究用試薬 ) < 目次 > 免疫組織染色手順 ( 前処理なし ) p2 免疫組織染色手順 ( マイクロウェーブ前処理 ) p3 免疫組織染色手順 ( オートクレーブ前処理 ) p4 免疫組織染色手順 ( トリプシン前処理 ) p5 免疫組織染色手順 ( ギ酸処理 ) p6 免疫
 < 目次 > 免疫組織染色手順 ( 前処理なし ) p2 免疫組織染色手順 ( マイクロウェーブ前処理 ) p3 免疫組織染色手順 ( オートクレーブ前処理 ) p4 免疫組織染色手順 ( トリプシン前処理 ) p5 免疫組織染色手順 ( ギ酸処理 ) p6 免疫") < 目次 > 免疫組織染色手順 ( 前処理なし ) p2 免疫組織染色手順 ( マイクロウェーブ前処理 ) p3 免疫組織染色手順 ( オートクレーブ前処理 ) p4 免疫組織染色手順 ( トリプシン前処理 ) p5 免疫組織染色手順 ( ギ酸処理 ) p6 免疫組織染色手順 ( ギ酸処理後 マイクロウェーブまたはオートクレーブ処理 )p7 抗原ペプチドによる抗体吸収試験 p8 ウエスタン ブロッティング
< 目次 > 免疫組織染色手順 ( 前処理なし ) p2 免疫組織染色手順 ( マイクロウェーブ前処理 ) p3 免疫組織染色手順 ( オートクレーブ前処理 ) p4 免疫組織染色手順 ( トリプシン前処理 ) p5 免疫組織染色手順 ( ギ酸処理 ) p6 免疫組織染色手順 ( ギ酸処理後 マイクロウェーブまたはオートクレーブ処理 )p7 抗原ペプチドによる抗体吸収試験 p8 ウエスタン ブロッティング
No. 1 Ⅰ. 緒言現在我国は超高齢社会を迎え それに伴い 高齢者の健康増進に関しては歯科医療も今後より重要な役割を担うことになる その中でも 部分欠損歯列の修復に伴う口腔内のメンテナンスがより一層必要となってきている しかし 高齢期は身体機能全般の変調を伴うことが多いため 口腔内環境の悪化を招く
 学位論文内容の要約 甲第 692 号論文提出者金野弘靖 論文題目 口腔レンサ球菌における環状ヌクレオチドの働きについて No. 1 Ⅰ. 緒言現在我国は超高齢社会を迎え それに伴い 高齢者の健康増進に関しては歯科医療も今後より重要な役割を担うことになる その中でも 部分欠損歯列の修復に伴う口腔内のメンテナンスがより一層必要となってきている しかし 高齢期は身体機能全般の変調を伴うことが多いため 口腔内環境の悪化を招くことに成り易い
学位論文内容の要約 甲第 692 号論文提出者金野弘靖 論文題目 口腔レンサ球菌における環状ヌクレオチドの働きについて No. 1 Ⅰ. 緒言現在我国は超高齢社会を迎え それに伴い 高齢者の健康増進に関しては歯科医療も今後より重要な役割を担うことになる その中でも 部分欠損歯列の修復に伴う口腔内のメンテナンスがより一層必要となってきている しかし 高齢期は身体機能全般の変調を伴うことが多いため 口腔内環境の悪化を招くことに成り易い
DNA/RNA調製法 実験ガイド
 DNA/RNA 調製法実験ガイド PCR の鋳型となる DNA を調製するにはいくつかの方法があり 検体の種類や実験目的に応じて適切な方法を選択します この文書では これらの方法について実際の操作方法を具体的に解説します また RNA 調製の際の注意事項や RNA 調製用のキット等をご紹介します - 目次 - 1 実験に必要なもの 2 コロニーからの DNA 調製 3 増菌培養液からの DNA 調製
DNA/RNA 調製法実験ガイド PCR の鋳型となる DNA を調製するにはいくつかの方法があり 検体の種類や実験目的に応じて適切な方法を選択します この文書では これらの方法について実際の操作方法を具体的に解説します また RNA 調製の際の注意事項や RNA 調製用のキット等をご紹介します - 目次 - 1 実験に必要なもの 2 コロニーからの DNA 調製 3 増菌培養液からの DNA 調製
無細胞タンパク質合成試薬キット Transdirect insect cell
 発現プラスミドの構築 1. インサート DNA の調製開始コドンは出来るだけ 5'UTR に近い位置に挿入して下さい 経験的に ptd1 の EcoRV/KpnI サイトへのライゲーション効率が最も高いことを確認しています 本プロトコルに従うと インサートサイズにも依りますが 90% 以上のコロニーがインサートの挿入されたクローンとして得られます 可能な限り EcoRV/KpnI サイトへ挿入されるお奨めします
発現プラスミドの構築 1. インサート DNA の調製開始コドンは出来るだけ 5'UTR に近い位置に挿入して下さい 経験的に ptd1 の EcoRV/KpnI サイトへのライゲーション効率が最も高いことを確認しています 本プロトコルに従うと インサートサイズにも依りますが 90% 以上のコロニーがインサートの挿入されたクローンとして得られます 可能な限り EcoRV/KpnI サイトへ挿入されるお奨めします
手順 ) 1) プライマーの設計 発注変異導入部位がプライマーのほぼ中央になるようにする 可能であれば 制限酵素サイトができるようにすると確認が容易になる プライマーは 25-45mer で TM 値が 78 以上になるようにする Tm= (%GC)-675/N-%mismatch
 1) プライマーの設計 発注変異導入部位がプライマーのほぼ中央になるようにする 可能であれば 制限酵素サイトができるようにすると確認が容易になる プライマーは 25-45mer で TM 値が 78 以上になるようにする Tm= (%GC)-675/N-%mismatch") Mutagenesis 目的 ) 既存の遺伝子に PCR を利用して変異を導入する 1 点変異導入方法 ) Quik Change Site-Directed Mutagenesis Kit(Stratagene) のプロトコールを流用 http://www.stratagene.com/products/showproduct.aspx?pid=131 Kit 中では DNA polymerase
Mutagenesis 目的 ) 既存の遺伝子に PCR を利用して変異を導入する 1 点変異導入方法 ) Quik Change Site-Directed Mutagenesis Kit(Stratagene) のプロトコールを流用 http://www.stratagene.com/products/showproduct.aspx?pid=131 Kit 中では DNA polymerase
■リアルタイムPCR実践編
 リアルタイム PCR 実践編 - SYBR Green I によるリアルタイム RT-PCR - 1. プライマー設計 (1)Perfect Real Time サポートシステムを利用し 設計済みのものを購入する ヒト マウス ラットの RefSeq 配列の大部分については Perfect Real Time サポートシステムが利用できます 目的の遺伝子を検索して購入してください (2) カスタム設計サービスを利用する
リアルタイム PCR 実践編 - SYBR Green I によるリアルタイム RT-PCR - 1. プライマー設計 (1)Perfect Real Time サポートシステムを利用し 設計済みのものを購入する ヒト マウス ラットの RefSeq 配列の大部分については Perfect Real Time サポートシステムが利用できます 目的の遺伝子を検索して購入してください (2) カスタム設計サービスを利用する
BKL Kit (Blunting Kination Ligation Kit)
") 製品コード 6126/6127 BKL Kit (Blunting Kination Ligation Kit) 説明書 PCR 産物を平滑末端ベクターにクローニングする場合 使用するポリメラーゼ酵素の種類により 3' 末端に余分に付加された塩基を除去し さらに 5' 末端をリン酸化する必要があります 本製品は これらの一連の反応を簡便に短時間に行うためのキットです PCR 産物の末端平滑化とリン酸化を同時に行うことにより
製品コード 6126/6127 BKL Kit (Blunting Kination Ligation Kit) 説明書 PCR 産物を平滑末端ベクターにクローニングする場合 使用するポリメラーゼ酵素の種類により 3' 末端に余分に付加された塩基を除去し さらに 5' 末端をリン酸化する必要があります 本製品は これらの一連の反応を簡便に短時間に行うためのキットです PCR 産物の末端平滑化とリン酸化を同時に行うことにより
TaKaRa PCR Human Papillomavirus Typing Set
 研究用 TaKaRa PCR Human Papillomavirus Typing Set 説明書 v201703da 本製品は さまざまなタイプの Human Papillomavirus(HPV) において塩基配列の相同性の高い領域に設定したプライマーを consensus primer として用いることにより HPV の E6 と E7 を含む領域 (228 ~ 268 bp) を共通に PCR
研究用 TaKaRa PCR Human Papillomavirus Typing Set 説明書 v201703da 本製品は さまざまなタイプの Human Papillomavirus(HPV) において塩基配列の相同性の高い領域に設定したプライマーを consensus primer として用いることにより HPV の E6 と E7 を含む領域 (228 ~ 268 bp) を共通に PCR
ChIP Reagents マニュアル
 ChIP Reagents ~ クロマチン免疫沈降 (ChIP) 法用ストック溶液セット ~ マニュアル ( 第 1 版 ) Code No. 318-07131 NIPPON GENE CO., LTD. 目次 Ⅰ 製品説明 1 Ⅱ セット内容 1 Ⅲ 保存 2 Ⅳ 使用上の注意 2 Ⅴ 使用例 2 < 本品以外に必要な試薬 機器など> 2 1) 磁気ビーズと一次抗体の反応 3 2)-1 細胞の調製
ChIP Reagents ~ クロマチン免疫沈降 (ChIP) 法用ストック溶液セット ~ マニュアル ( 第 1 版 ) Code No. 318-07131 NIPPON GENE CO., LTD. 目次 Ⅰ 製品説明 1 Ⅱ セット内容 1 Ⅲ 保存 2 Ⅳ 使用上の注意 2 Ⅴ 使用例 2 < 本品以外に必要な試薬 機器など> 2 1) 磁気ビーズと一次抗体の反応 3 2)-1 細胞の調製
PowerPoint プレゼンテーション
 2013 年 11 月 20 日 ( 水 ) バイオ情報解析演習 ウェブツールを活用した生物情報解析 (4) 遺伝子のクローニング設計 有用物質生産菌を合理的に作ろう! 設計 試作 ベンチテスト 完成 プラスミド 効率的な代謝経路を設計する 文献調査代謝パスウェイの探索代謝シミュレーション 実際に微生物に組み込む データベースから有用遺伝子を探索する遺伝子組換え技術 培養をして問題点を突き止める 培養代謝物量
2013 年 11 月 20 日 ( 水 ) バイオ情報解析演習 ウェブツールを活用した生物情報解析 (4) 遺伝子のクローニング設計 有用物質生産菌を合理的に作ろう! 設計 試作 ベンチテスト 完成 プラスミド 効率的な代謝経路を設計する 文献調査代謝パスウェイの探索代謝シミュレーション 実際に微生物に組み込む データベースから有用遺伝子を探索する遺伝子組換え技術 培養をして問題点を突き止める 培養代謝物量
大学院博士課程共通科目ベーシックプログラム
 平成 30 年度医科学専攻共通科目 共通基礎科目実習 ( 旧コア実習 ) 概要 1 ). 大学院生が所属する教育研究分野における実習により単位認定可能な実習項目 ( コア実習項目 ) 1. 組換え DNA 技術実習 2. 生体物質の調製と解析実習 3. 薬理学実習 4. ウイルス学実習 5. 免疫学実習 6. 顕微鏡試料作成法実習 7. ゲノム医学実習 8. 共焦点レーザー顕微鏡実習 2 ). 実習を担当する教育研究分野においてのみ単位認定可能な実習項目
平成 30 年度医科学専攻共通科目 共通基礎科目実習 ( 旧コア実習 ) 概要 1 ). 大学院生が所属する教育研究分野における実習により単位認定可能な実習項目 ( コア実習項目 ) 1. 組換え DNA 技術実習 2. 生体物質の調製と解析実習 3. 薬理学実習 4. ウイルス学実習 5. 免疫学実習 6. 顕微鏡試料作成法実習 7. ゲノム医学実習 8. 共焦点レーザー顕微鏡実習 2 ). 実習を担当する教育研究分野においてのみ単位認定可能な実習項目
PrimeSTAR® Mutagenesis Basal Kit
 PrimeSTAR Mutagenesis Basal Kit 説明書 v201107da 目次 I. キットの内容...3 II. 保存...3 III. 本システムの原理...3 IV. プライマー設計について...5 V. 操作...8 VI. 実験例...10 VII. トラブルシューティング...13 VIII. 関連製品...13 IX. 注意...13 2 PrimeSTAR Mutagenesis
PrimeSTAR Mutagenesis Basal Kit 説明書 v201107da 目次 I. キットの内容...3 II. 保存...3 III. 本システムの原理...3 IV. プライマー設計について...5 V. 操作...8 VI. 実験例...10 VII. トラブルシューティング...13 VIII. 関連製品...13 IX. 注意...13 2 PrimeSTAR Mutagenesis
Microsoft PowerPoint - 雑誌会 pptx
 石井研雑誌会 No. 1148 2014. 1. 17 新井博之 Neisseria gonorrhoeae における cbb 3 oxidase の CcoP サブユニットと cytochrome c 2 を介した微好気呼吸と脱窒のリンク 背景 1 Neisseria gonorrhoeae (gocococcus, 淋菌 ) -proteobacteria の病原性菌末端酸化酵素は cbb 3
石井研雑誌会 No. 1148 2014. 1. 17 新井博之 Neisseria gonorrhoeae における cbb 3 oxidase の CcoP サブユニットと cytochrome c 2 を介した微好気呼吸と脱窒のリンク 背景 1 Neisseria gonorrhoeae (gocococcus, 淋菌 ) -proteobacteria の病原性菌末端酸化酵素は cbb 3
Microsoft Word - FMB_Text(PCR) _ver3.doc
 _ver3.doc") 2.PCR 法による DNA の増幅 現代の分子生物学において その進歩に最も貢献した実験法の1つが PCR(Polymerase chain reaction) 法である PCR 法は極めて微量の DNA サンプルから特定の DNA 断片を短時間に大量に増幅することができる方法であり 多大な時間と労力を要した遺伝子クローニングを過去のものとしてしまった また その操作の簡便さから 現在では基礎研究のみならず臨床遺伝子診断から食品衛生検査
2.PCR 法による DNA の増幅 現代の分子生物学において その進歩に最も貢献した実験法の1つが PCR(Polymerase chain reaction) 法である PCR 法は極めて微量の DNA サンプルから特定の DNA 断片を短時間に大量に増幅することができる方法であり 多大な時間と労力を要した遺伝子クローニングを過去のものとしてしまった また その操作の簡便さから 現在では基礎研究のみならず臨床遺伝子診断から食品衛生検査
cDNA cloning by PCR
 cdna cloning/subcloning by PCR 1. 概要 2014. 4 ver.1 by A. Goto & K. Takeda 一般的に 細胞または組織由来の RNA から作製した cdna(cdna pool) から 特定の cdna をベクターに組み込む操作を cdna cloning と呼ぶ その際 制限酵素認識配列を付与したオリゴ DNA primer を用いた PCR
cdna cloning/subcloning by PCR 1. 概要 2014. 4 ver.1 by A. Goto & K. Takeda 一般的に 細胞または組織由来の RNA から作製した cdna(cdna pool) から 特定の cdna をベクターに組み込む操作を cdna cloning と呼ぶ その際 制限酵素認識配列を付与したオリゴ DNA primer を用いた PCR
論文題目 腸管分化に関わるmiRNAの探索とその発現制御解析
 論文題目 腸管分化に関わる microrna の探索とその発現制御解析 氏名日野公洋 1. 序論 microrna(mirna) とは細胞内在性の 21 塩基程度の機能性 RNA のことであり 部分的相補的な塩基認識を介して標的 RNA の翻訳抑制や不安定化を引き起こすことが知られている mirna は細胞分化や増殖 ガン化やアポトーシスなどに関与していることが報告されており これら以外にも様々な細胞諸現象に関与していると考えられている
論文題目 腸管分化に関わる microrna の探索とその発現制御解析 氏名日野公洋 1. 序論 microrna(mirna) とは細胞内在性の 21 塩基程度の機能性 RNA のことであり 部分的相補的な塩基認識を介して標的 RNA の翻訳抑制や不安定化を引き起こすことが知られている mirna は細胞分化や増殖 ガン化やアポトーシスなどに関与していることが報告されており これら以外にも様々な細胞諸現象に関与していると考えられている
TaKaRa PCR Human Papillomavirus Detection Set
 研究用 TaKaRa PCR Human Papillomavirus Detection Set 説明書 v201703da 本製品は子宮頸癌において最も高頻度に検出される Human Papillomavirus(HPV)16 18 および 33 型をそれぞれ特異的に増幅し 検出するセットです HPV16 18 および 33 型の E6 を含む領域 (140 bp ただし HPV33 型は 141
研究用 TaKaRa PCR Human Papillomavirus Detection Set 説明書 v201703da 本製品は子宮頸癌において最も高頻度に検出される Human Papillomavirus(HPV)16 18 および 33 型をそれぞれ特異的に増幅し 検出するセットです HPV16 18 および 33 型の E6 を含む領域 (140 bp ただし HPV33 型は 141
研究目的 1. 電波ばく露による免疫細胞への影響に関する研究 我々の体には 恒常性を保つために 生体内に侵入した異物を生体外に排除する 免疫と呼ばれる防御システムが存在する 免疫力の低下は感染を引き起こしやすくなり 健康を損ないやすくなる そこで 2 10W/kgのSARで電波ばく露を行い 免疫細胞
 資料 - 生電 6-3 免疫細胞及び神経膠細胞を対象としたマイクロ波照射影響に関する実験評価 京都大学首都大学東京 宮越順二 成田英二郎 櫻井智徳多氣昌生 鈴木敏久 日 : 平成 23 年 7 月 22 日 ( 金 ) 場所 : 総務省第 1 特別会議室 研究目的 1. 電波ばく露による免疫細胞への影響に関する研究 我々の体には 恒常性を保つために 生体内に侵入した異物を生体外に排除する 免疫と呼ばれる防御システムが存在する
資料 - 生電 6-3 免疫細胞及び神経膠細胞を対象としたマイクロ波照射影響に関する実験評価 京都大学首都大学東京 宮越順二 成田英二郎 櫻井智徳多氣昌生 鈴木敏久 日 : 平成 23 年 7 月 22 日 ( 金 ) 場所 : 総務省第 1 特別会議室 研究目的 1. 電波ばく露による免疫細胞への影響に関する研究 我々の体には 恒常性を保つために 生体内に侵入した異物を生体外に排除する 免疫と呼ばれる防御システムが存在する
Western BLoT Rapid Detect
 研究用 Western BLoT Rapid Detect 説明書 v201212 Western BLoT Rapid Detect は 標識二次抗体の代わりに独自の IgG Detector(HRP labeled) を利用して一次抗体を検出するウェスタンブロッティング専用の検出試薬キットです 本製品を利用することで 標識二次抗体を用いて検出する従来法ではできなかった迅速検出 高感度検出 シグナルの増強
研究用 Western BLoT Rapid Detect 説明書 v201212 Western BLoT Rapid Detect は 標識二次抗体の代わりに独自の IgG Detector(HRP labeled) を利用して一次抗体を検出するウェスタンブロッティング専用の検出試薬キットです 本製品を利用することで 標識二次抗体を用いて検出する従来法ではできなかった迅速検出 高感度検出 シグナルの増強
製品コード 3372 研究用 pcold GST DNA 説明書 v201909da
 研究用 pcold GST DNA 説明書 v201909da タンパク質の構造や機能の解明はポストゲノムの重要な研究対象であり 効率の良いタンパク質生産システムはポストゲノム解析に必須の基盤技術です 組換えタンパク質の生産には大腸菌を宿主とする発現系が広く利用されています しかしながら 大腸菌発現系は扱いやすく低コストである反面 遺伝子によっては発現できない あるいは発現タンパク質が不溶化するという問題が起こることがあります
研究用 pcold GST DNA 説明書 v201909da タンパク質の構造や機能の解明はポストゲノムの重要な研究対象であり 効率の良いタンパク質生産システムはポストゲノム解析に必須の基盤技術です 組換えタンパク質の生産には大腸菌を宿主とする発現系が広く利用されています しかしながら 大腸菌発現系は扱いやすく低コストである反面 遺伝子によっては発現できない あるいは発現タンパク質が不溶化するという問題が起こることがあります
Human Cell-Free Protein Expression System
 研究用 Human Cell-Free Protein Expression System 説明書 v201807da Human Cell-Free Protein Expression System は ヒト細胞株由来の細胞抽出液を利用した無細胞タンパク質合成システムです 本システムの Cell Lysate には in vitro でのタンパク質合成反応に必要な各種因子 ( リボソーム 翻訳開始
研究用 Human Cell-Free Protein Expression System 説明書 v201807da Human Cell-Free Protein Expression System は ヒト細胞株由来の細胞抽出液を利用した無細胞タンパク質合成システムです 本システムの Cell Lysate には in vitro でのタンパク質合成反応に必要な各種因子 ( リボソーム 翻訳開始
DNA 抽出条件かき取った花粉 1~3 粒程度を 3 μl の抽出液 (10 mm Tris/HCl [ph8.0] 10 mm EDTA 0.01% SDS 0.2 mg/ml Proteinase K) に懸濁し 37 C 60 min そして 95 C 10 min の処理を行うことで DNA
![DNA 抽出条件かき取った花粉 1~3 粒程度を 3 μl の抽出液 (10 mm Tris/HCl [ph8.0] 10 mm EDTA 0.01% SDS 0.2 mg/ml Proteinase K) に懸濁し 37 C 60 min そして 95 C 10 min の処理を行うことで DNA](/thumbs/91/104738989.jpg "DNA 抽出条件かき取った花粉 1~3 粒程度を 3 μl の抽出液 (10 mm Tris/HCl [ph8.0] 10 mm EDTA 0.01% SDS 0.2 mg/ml Proteinase K) に懸濁し 37 C 60 min そして 95 C 10 min の処理を行うことで DNA") 組換えイネ花粉の飛散試験 交雑試験 1. 飛散試験 目的 隔離圃場内の試験区で栽培している組換えイネ S-C 系統 及び AS-D 系統の開花時における花粉の飛散状況を確認するため 方法 (1) H23 年度は 7 月末からの低温の影響を受け例年の開花時期よりも遅れ 試験に用いた組換えイネの開花が最初に確認されたのは S-C 系統 及び AS-D 系統ともに 8 月 13 日であった そこで予め準備しておいた花粉トラップ
組換えイネ花粉の飛散試験 交雑試験 1. 飛散試験 目的 隔離圃場内の試験区で栽培している組換えイネ S-C 系統 及び AS-D 系統の開花時における花粉の飛散状況を確認するため 方法 (1) H23 年度は 7 月末からの低温の影響を受け例年の開花時期よりも遅れ 試験に用いた組換えイネの開花が最初に確認されたのは S-C 系統 及び AS-D 系統ともに 8 月 13 日であった そこで予め準備しておいた花粉トラップ
Untitled
 上原記念生命科学財団研究報告集, 25 (2011) 86. 線虫 C. elegans およびマウスをモデル動物とした体細胞レベルで生じる性差の解析 井上英樹 Key words: 性差, ストレス応答,DMRT 立命館大学生命科学部生命医科学科 緒言性差は雌雄の性に分かれた動物にみられ, 生殖能力の違いだけでなく形態, 行動などそれぞれの性の間でみられる様々な差異と定義される. 性差は, 形態や行動だけでなく疾患の発症リスクの男女差といった生理的なレベルの差異も含まれる.
上原記念生命科学財団研究報告集, 25 (2011) 86. 線虫 C. elegans およびマウスをモデル動物とした体細胞レベルで生じる性差の解析 井上英樹 Key words: 性差, ストレス応答,DMRT 立命館大学生命科学部生命医科学科 緒言性差は雌雄の性に分かれた動物にみられ, 生殖能力の違いだけでなく形態, 行動などそれぞれの性の間でみられる様々な差異と定義される. 性差は, 形態や行動だけでなく疾患の発症リスクの男女差といった生理的なレベルの差異も含まれる.
Microsoft Word - タンパク質溶液内酵素消化 Thermo
 タンパク質の溶液内酵素溶液内酵素消化 ( 質量分析用サンプル調製 ) 質量分析計によるタンパク質解析においては 一般的にタンパク質を還元 アルキル化した後にトリプシン等で酵素消化して得られた消化ペプチドサンプルが用いられます 本資料ではこのサンプル調製について 専用キットを用いて行う方法 各種試薬や酵素を用いて行う方法 また関連情報として タンパク質の定量法についてご紹介しています 内容 1 培養細胞の酵素消化
タンパク質の溶液内酵素溶液内酵素消化 ( 質量分析用サンプル調製 ) 質量分析計によるタンパク質解析においては 一般的にタンパク質を還元 アルキル化した後にトリプシン等で酵素消化して得られた消化ペプチドサンプルが用いられます 本資料ではこのサンプル調製について 専用キットを用いて行う方法 各種試薬や酵素を用いて行う方法 また関連情報として タンパク質の定量法についてご紹介しています 内容 1 培養細胞の酵素消化
Taro-kv12250.jtd
 ニューカッスル病 マレック病 ( ニューカッスル病ウイルス由来 F 蛋白遺伝子導入マレック病ウイルス 1 型 ) 凍結生ワクチン 平成 22 年 8 月 12 日 ( 告示第 2288 号 ) 新規追加 ニューカッスル病ウイルスのF 蛋白をコードする遺伝子を弱毒マレック病ウイルス (1 型 ) に挿入して得られた組換え体ウイルスを培養細胞で増殖させて得た感染細胞浮遊液を凍結したワクチンである 1 小分製品の試験
ニューカッスル病 マレック病 ( ニューカッスル病ウイルス由来 F 蛋白遺伝子導入マレック病ウイルス 1 型 ) 凍結生ワクチン 平成 22 年 8 月 12 日 ( 告示第 2288 号 ) 新規追加 ニューカッスル病ウイルスのF 蛋白をコードする遺伝子を弱毒マレック病ウイルス (1 型 ) に挿入して得られた組換え体ウイルスを培養細胞で増殖させて得た感染細胞浮遊液を凍結したワクチンである 1 小分製品の試験
組織からのゲノム DNA 抽出キット Tissue Genomic DNA Extraction Mini Kit 目次基本データ 3 キットの内容 3 重要事項 4 操作 4 サンプル別プロトコール 7 トラブルシューティング 9 * 本製品は研究用です *
 組織からのゲノム DNA 抽出キット Tissue Genomic DNA Extraction Mini Kit 目次基本データ 3 キットの内容 3 重要事項 4 操作 4 サンプル別プロトコール 7 トラブルシューティング 9 * 本製品は研究用です * 2 本キットは動物の組織からトータル DNA を迅速に効率よく抽出するようにデザインされています また 細菌 固定組織 酵母用のプロトコールも用意しています
組織からのゲノム DNA 抽出キット Tissue Genomic DNA Extraction Mini Kit 目次基本データ 3 キットの内容 3 重要事項 4 操作 4 サンプル別プロトコール 7 トラブルシューティング 9 * 本製品は研究用です * 2 本キットは動物の組織からトータル DNA を迅速に効率よく抽出するようにデザインされています また 細菌 固定組織 酵母用のプロトコールも用意しています
<4D F736F F D F4390B38CE3816A90528DB88C8B89CA2E646F63>
 学位論文の内容の要旨 論文提出者氏名 論文審査担当者 論文題目 主査 荒川真一 御給美沙 副査木下淳博横山三紀 Thrombospondin-1 Production is Enhanced by Porphyromonas gingivalis Lipopolysaccharide in THP-1 Cells ( 論文の内容の要旨 ) < 要旨 > 歯周炎はグラム陰性嫌気性細菌によって引き起こされる慢性炎症性疾患であり
学位論文の内容の要旨 論文提出者氏名 論文審査担当者 論文題目 主査 荒川真一 御給美沙 副査木下淳博横山三紀 Thrombospondin-1 Production is Enhanced by Porphyromonas gingivalis Lipopolysaccharide in THP-1 Cells ( 論文の内容の要旨 ) < 要旨 > 歯周炎はグラム陰性嫌気性細菌によって引き起こされる慢性炎症性疾患であり
Taro-SV02500b.jtd
 牛大腸菌性下痢症 (K99 保有全菌体 FY 保有全菌体 31A 保有全菌体 O78 全菌体 )( アジュバント加 ) 不活化ワクチン 平成 21 年 11 月 12 日 ( 告示第 1569 号 ) 一部改正 1 定義線毛抗原 K99 FY 及び 31A を保有する大腸菌並びに O78 の大腸菌の培養菌液を不活化したものを混合し アルミニウムゲルアジュバントを添加したワクチンである 2 製法 2.1
牛大腸菌性下痢症 (K99 保有全菌体 FY 保有全菌体 31A 保有全菌体 O78 全菌体 )( アジュバント加 ) 不活化ワクチン 平成 21 年 11 月 12 日 ( 告示第 1569 号 ) 一部改正 1 定義線毛抗原 K99 FY 及び 31A を保有する大腸菌並びに O78 の大腸菌の培養菌液を不活化したものを混合し アルミニウムゲルアジュバントを添加したワクチンである 2 製法 2.1
大学院委員会について
 博士学位請求論文 審査報告書 審査委員 ( 主査 ) 農学部専任准教授村上周一朗 ( 副査 ) 農学部専任教授中島春紫 ( 副査 ) 農学部専任准教授前田理久 2014 年 1 月 17 日 1 論文提出者氏名高橋結 2 論文題名 ( 邦文題 ) ハイイロジェントルキツネザルの糞由来 Aspergillus niger E-1 株の有する xylanase に関する研究 ( 欧文訳 )Study on
博士学位請求論文 審査報告書 審査委員 ( 主査 ) 農学部専任准教授村上周一朗 ( 副査 ) 農学部専任教授中島春紫 ( 副査 ) 農学部専任准教授前田理久 2014 年 1 月 17 日 1 論文提出者氏名高橋結 2 論文題名 ( 邦文題 ) ハイイロジェントルキツネザルの糞由来 Aspergillus niger E-1 株の有する xylanase に関する研究 ( 欧文訳 )Study on
A, B 2 B 5 A C A ADP A G ADP ADP G camp B GM A B CTB Tochikubo et al., 1998 CTB CTB Dertzbaugh & Elson, 1993a; Yu et al., 1994; Shi et al., 1995 CTB C
 2004 Analysis of the expression of cholera toxin B-subunit gene 1065010 2004 8 26 1 A, B 2 B 5 A C A ADP A G ADP ADP G camp B GM A B CTB Tochikubo et al., 1998 CTB CTB Dertzbaugh & Elson, 1993a; Yu et
2004 Analysis of the expression of cholera toxin B-subunit gene 1065010 2004 8 26 1 A, B 2 B 5 A C A ADP A G ADP ADP G camp B GM A B CTB Tochikubo et al., 1998 CTB CTB Dertzbaugh & Elson, 1993a; Yu et
DNA Blunting Kit
 研究用 DNA Blunting Kit 説明書 v201508da DNA Blunting Kit は T4 DNA Polymerase の 5' 3' polymerase 活性と 3' 5' exonuclease 活性を利用して DNA の末端を平滑化することにより 突出末端の DNA でも簡単に平滑末端のベクターにライゲーションできるシステムです DNA が 5' 突出末端の場合は 5'
研究用 DNA Blunting Kit 説明書 v201508da DNA Blunting Kit は T4 DNA Polymerase の 5' 3' polymerase 活性と 3' 5' exonuclease 活性を利用して DNA の末端を平滑化することにより 突出末端の DNA でも簡単に平滑末端のベクターにライゲーションできるシステムです DNA が 5' 突出末端の場合は 5'
Microsoft Word - H18年度生命工学報告書 Astro ver.3
 1. Rotavirus Norovirus Adenovirus Astrovirus Sapovirus 5 1) Human Astrovirus HastV 1975 Madeley Cosgrove EM RNA 30nm 5 6 Fig. 1 2) HastV 8 3 5 HastV ELISA Enzyme-Linked ImmunoSorbent Assay RIA Radio immunoassay
1. Rotavirus Norovirus Adenovirus Astrovirus Sapovirus 5 1) Human Astrovirus HastV 1975 Madeley Cosgrove EM RNA 30nm 5 6 Fig. 1 2) HastV 8 3 5 HastV ELISA Enzyme-Linked ImmunoSorbent Assay RIA Radio immunoassay
( 様式甲 5) 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 教授 大道正英 髙橋優子 副査副査 教授教授 岡 田 仁 克 辻 求 副査 教授 瀧内比呂也 主論文題名 Versican G1 and G3 domains are upregulated and latent trans
 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 教授 大道正英 髙橋優子 副査副査 教授教授 岡 田 仁 克 辻 求 副査 教授 瀧内比呂也 主論文題名 Versican G1 and G3 domains are upregulated and latent trans") ( 様式甲 5) 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 大道正英 髙橋優子 副査副査 岡 田 仁 克 辻 求 副査 瀧内比呂也 主論文題名 Versican G1 and G3 domains are upregulated and latent transforming growth factor- binding protein-4 is downregulated in breast
( 様式甲 5) 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 大道正英 髙橋優子 副査副査 岡 田 仁 克 辻 求 副査 瀧内比呂也 主論文題名 Versican G1 and G3 domains are upregulated and latent transforming growth factor- binding protein-4 is downregulated in breast
Microsoft Word - Dr. Abe.doc
 3 ステップ アビジン - ビオチンシステム (SAB 法 ) とポリマー法 慶應義塾大学医学部病理学教室阿部仁 はじめに 免疫組織化学は Coons らが蛍光色素を抗体に標識した蛍光抗体法の技術を確立してから Singers のフェリチン抗体法を経て 1967 年に Nakane と Pierce により標識物質に西洋ワサビペルオキシダーゼ (horseradish peroxidase:hrp)
3 ステップ アビジン - ビオチンシステム (SAB 法 ) とポリマー法 慶應義塾大学医学部病理学教室阿部仁 はじめに 免疫組織化学は Coons らが蛍光色素を抗体に標識した蛍光抗体法の技術を確立してから Singers のフェリチン抗体法を経て 1967 年に Nakane と Pierce により標識物質に西洋ワサビペルオキシダーゼ (horseradish peroxidase:hrp)
プロトコル 細胞 増殖 / 毒性酸化ストレス分子生物学細胞内蛍光プローブ細胞染色ミトコンドリア関連試薬細菌研究用試薬膜タンパク質可溶化剤ラベル化剤二価性試薬イオン電極 その他 機能性有機材料 酵素 (POD,ALP) を標識したい 利用製品 < 少量抗体 (10μg) 標識用 > Ab-10 Rap
 を標識したい 利用製品 < 少量抗体 (10μg) 標識用 > Ab-10 Rap") 増殖 / 毒性酸化ストレス分子生物学内蛍光プローブ染色ミトコンドリア関連試薬細菌研究用試薬膜タンパク質可溶化剤ラベル化剤二価性試薬イオン電極 機能性有機材料 酵素 (PD,ALP) を標識したい 利用製品 < 少量抗体 (0μg) 標識用 > Ab-0 Rapid Peroxidase Labeling Kit < 抗体 タンパク質 (0-00μg) 標識用 > Peroxidase Labeling
増殖 / 毒性酸化ストレス分子生物学内蛍光プローブ染色ミトコンドリア関連試薬細菌研究用試薬膜タンパク質可溶化剤ラベル化剤二価性試薬イオン電極 機能性有機材料 酵素 (PD,ALP) を標識したい 利用製品 < 少量抗体 (0μg) 標識用 > Ab-0 Rapid Peroxidase Labeling Kit < 抗体 タンパク質 (0-00μg) 標識用 > Peroxidase Labeling
ISOSPIN Plasmid
 プラスミド DNA 抽出キット ISOSPIN Plasmid マニュアル ( 第 5 版 ) Code No. 318-07991 NIPPON GENE CO., LTD. I 製品説明 ISOSPIN Plasmid( アイソスピンプラスミド ) は スピンカラムを用いて簡単に大腸菌から高純度なプラスミド DNA を抽出できるキットです 本キットは カオトロピックイオン存在下で DNA がシリカへ吸着する原理を応用しており
プラスミド DNA 抽出キット ISOSPIN Plasmid マニュアル ( 第 5 版 ) Code No. 318-07991 NIPPON GENE CO., LTD. I 製品説明 ISOSPIN Plasmid( アイソスピンプラスミド ) は スピンカラムを用いて簡単に大腸菌から高純度なプラスミド DNA を抽出できるキットです 本キットは カオトロピックイオン存在下で DNA がシリカへ吸着する原理を応用しており
Untitled
 上原記念生命科学財団研究報告集, 23(2009) 149. サルエイズウイルスのヒトへの感染伝播を規定する宿主制御因子の解明 武内寛明 Key words: エイズウイルス, 異種間感染, 感染症, 人畜共通感染症, 新興感染症 東京大学医科学研究所感染症国際研究センター微生物学分野 緒言ヒト後天性免疫不全症候群 ( ヒトエイズ ) は, ヒト免疫不全ウイルス (HIV) によって引き起こされる慢性持続感染症である.
上原記念生命科学財団研究報告集, 23(2009) 149. サルエイズウイルスのヒトへの感染伝播を規定する宿主制御因子の解明 武内寛明 Key words: エイズウイルス, 異種間感染, 感染症, 人畜共通感染症, 新興感染症 東京大学医科学研究所感染症国際研究センター微生物学分野 緒言ヒト後天性免疫不全症候群 ( ヒトエイズ ) は, ヒト免疫不全ウイルス (HIV) によって引き起こされる慢性持続感染症である.
SPP System Set,SPP System I,SPP System II,SPP System III,SPP System IV
 研究用 SPP System Set ( 製品コード 3366) SPP System I ( 製品コード 3367) SPP System II ( 製品コード 3368) SPP System III ( 製品コード 3369) SPP System IV ( 製品コード 3370) 説明書 v201612da 目次 I. 製品説明...3 II. 内容...4 III. 保存...4 IV.
研究用 SPP System Set ( 製品コード 3366) SPP System I ( 製品コード 3367) SPP System II ( 製品コード 3368) SPP System III ( 製品コード 3369) SPP System IV ( 製品コード 3370) 説明書 v201612da 目次 I. 製品説明...3 II. 内容...4 III. 保存...4 IV.
PanaceaGel ゲル内細胞の観察 解析方法 1. ゲル内細胞の免疫染色 蛍光観察の方法 以下の 1-1, 1-2 に関して ゲルをスパーテルなどで取り出す際は 4% パラホルムアルデヒドで固定してから行うとゲルを比較的簡単に ( 壊さずに ) 取り出すことが可能です セルカルチャーインサートを
 取り出すことが可能です セルカルチャーインサートを") 1. ゲル内細胞の免疫染色 蛍光観察の方法 以下の 1-1, 1-2 に関して ゲルをスパーテルなどで取り出す際は 4% パラホルムアルデヒドで固定してから行うとゲルを比較的簡単に ( 壊さずに ) 取り出すことが可能です セルカルチャーインサートを用いた培養ではインサート底面をメス等で切り取ることで またウェルプレートを用いた培養ではピペットの水流でゲル ( 固定後 ) を底面から浮かすことで 回収しやすくなります
1. ゲル内細胞の免疫染色 蛍光観察の方法 以下の 1-1, 1-2 に関して ゲルをスパーテルなどで取り出す際は 4% パラホルムアルデヒドで固定してから行うとゲルを比較的簡単に ( 壊さずに ) 取り出すことが可能です セルカルチャーインサートを用いた培養ではインサート底面をメス等で切り取ることで またウェルプレートを用いた培養ではピペットの水流でゲル ( 固定後 ) を底面から浮かすことで 回収しやすくなります
Bacterial 16S rDNA PCR Kit
 研究用 Bacterial 16S rdna PCR Kit 説明書 v201802da 微生物の同定は 形態的特徴 生理 生化学的性状 化学分類学的性状などを利用して行われますが これらの方法では同定までに時間を要します また 同定が困難な場合や正しい結果が得られない場合もあります 近年 微生物同定にも分子生物学を利用した方法が採用されるようになり 微生物の持つ DNA を対象として解析を行う方法が活用されています
研究用 Bacterial 16S rdna PCR Kit 説明書 v201802da 微生物の同定は 形態的特徴 生理 生化学的性状 化学分類学的性状などを利用して行われますが これらの方法では同定までに時間を要します また 同定が困難な場合や正しい結果が得られない場合もあります 近年 微生物同定にも分子生物学を利用した方法が採用されるようになり 微生物の持つ DNA を対象として解析を行う方法が活用されています
 4-4 in situ PCR 研究室 in situ PCR 標的菌種と遺伝子 ( 報告年 ) 応用 Dept. of Marine Sciences University of Georgia USA Escherichia coli O157slt II Escherichia coli 16SrDNA Ralstonia eutropha phl (1997,1998,1999,2002)
4-4 in situ PCR 研究室 in situ PCR 標的菌種と遺伝子 ( 報告年 ) 応用 Dept. of Marine Sciences University of Georgia USA Escherichia coli O157slt II Escherichia coli 16SrDNA Ralstonia eutropha phl (1997,1998,1999,2002)
Untitled
 上原記念生命科学財団研究報告集, 26 (2012) 75. 哺乳類のゴルジ体ストレス応答の分子機構の解明 吉田秀郎 Key words: ゴルジ体, 小胞体, 転写, ストレス応答, 細胞小器官 兵庫県立大学大学院生命理学研究科生体物質化学 Ⅱ 講座 緒言細胞内には様々な細胞小器官が存在して細胞の機能を分担しているが, その存在量は細胞の需要に応じて厳密に制御されており, 必要な時に必要な細胞小器官が必要な量だけ増強される.
上原記念生命科学財団研究報告集, 26 (2012) 75. 哺乳類のゴルジ体ストレス応答の分子機構の解明 吉田秀郎 Key words: ゴルジ体, 小胞体, 転写, ストレス応答, 細胞小器官 兵庫県立大学大学院生命理学研究科生体物質化学 Ⅱ 講座 緒言細胞内には様々な細胞小器官が存在して細胞の機能を分担しているが, その存在量は細胞の需要に応じて厳密に制御されており, 必要な時に必要な細胞小器官が必要な量だけ増強される.
37-4.indd
 ISSN 0916-3328 東京大学アイソトープ総合センター VOL. 37 NO. 4 2007. 3. 26 アイソトープ総合センターの将来展望 40 18 18 20 3 800 1/3 1300 14 18 20 3 19 2 生きた細胞内の分子過程を 時間 空間計測する蛍光プローブの開発 1. はじめに Fura-2 Fura-2 198 1, 2 3, 4 2. 生体脂質分子の機能とその分析
ISSN 0916-3328 東京大学アイソトープ総合センター VOL. 37 NO. 4 2007. 3. 26 アイソトープ総合センターの将来展望 40 18 18 20 3 800 1/3 1300 14 18 20 3 19 2 生きた細胞内の分子過程を 時間 空間計測する蛍光プローブの開発 1. はじめに Fura-2 Fura-2 198 1, 2 3, 4 2. 生体脂質分子の機能とその分析
TaKaRa PCR FLT3/ITD Mutation Detection Set
 研究用 TaKaRa PCR FLT3/ITD Mutation Detection Set 説明書 v201706da 本製品は FLT3 (FMS-like tyrosine kinase 3) 遺伝子の JM(Juxtamembrane) 領域周辺に起こる Internal Tandem Duplication(ITD) 変異の有無を検出するためのセットです FLT3 遺伝子の ITD 変異は
研究用 TaKaRa PCR FLT3/ITD Mutation Detection Set 説明書 v201706da 本製品は FLT3 (FMS-like tyrosine kinase 3) 遺伝子の JM(Juxtamembrane) 領域周辺に起こる Internal Tandem Duplication(ITD) 変異の有無を検出するためのセットです FLT3 遺伝子の ITD 変異は
p _04本文
 l l l l l l 4 浦上財団研究報告書 Vol.15 2007 では 35Kおよび16K蛋白が各々70K 34Kの2量 3. 3 陰イオン交換クロマトグラフィー 体蛋白となって溶出していた SDS-PAGEは強い 陰極側採取液の35K 16K蛋白および通電前溶 還元状態で実施しているため これらの2量体は 液から得られた18K蛋白画分を陰イオン交換クロ ジスルフィド結合によって形成されたものでない
l l l l l l 4 浦上財団研究報告書 Vol.15 2007 では 35Kおよび16K蛋白が各々70K 34Kの2量 3. 3 陰イオン交換クロマトグラフィー 体蛋白となって溶出していた SDS-PAGEは強い 陰極側採取液の35K 16K蛋白および通電前溶 還元状態で実施しているため これらの2量体は 液から得られた18K蛋白画分を陰イオン交換クロ ジスルフィド結合によって形成されたものでない
PowerPoint プレゼンテーション
 植物 DA の抽出と増幅 1 背景 植物種子から DA 抽出する際 有機溶媒や酵素を用いた処理 遠心分離装置を使った操作など 煩雑で時間のかかる工程が用いられてきた カ技ネ術カの 検体採取 簡易 DA 抽出キット version2( 抽出キット ) と高速増幅用 DA Polymerase ( 増幅キット ) を組合わせる事で 遺伝子検査時間を短縮可能! 抽出キット * 抽出 DA を 増幅キット
植物 DA の抽出と増幅 1 背景 植物種子から DA 抽出する際 有機溶媒や酵素を用いた処理 遠心分離装置を使った操作など 煩雑で時間のかかる工程が用いられてきた カ技ネ術カの 検体採取 簡易 DA 抽出キット version2( 抽出キット ) と高速増幅用 DA Polymerase ( 増幅キット ) を組合わせる事で 遺伝子検査時間を短縮可能! 抽出キット * 抽出 DA を 増幅キット
鶏大腸菌症 ( 組換え型 F11 線毛抗原 ベロ細胞毒性抗原 )( 油性アジュバント加 ) 不活化ワクチン平成 20 年 6 月 6 日 ( 告示第 913 号 ) 新規追加 平成 25 年 9 月 26 日 ( 告示第 2480 号 ) 一部改正 1 定義組換え型 F11 線毛抗原産生大腸菌及びベ
( 油性アジュバント加 ) 不活化ワクチン平成 20 年 6 月 6 日 ( 告示第 913 号 ) 新規追加 平成 25 年 9 月 26 日 ( 告示第 2480 号 ) 一部改正 1 定義組換え型 F11 線毛抗原産生大腸菌及びベ") 鶏大腸菌症 ( 組換え型 F11 線毛抗原 ベロ細胞毒性抗原 )( 油性アジュバント加 ) 不活化ワクチン平成 20 年 6 月 6 日 ( 告示第 913 号 ) 新規追加 平成 25 年 9 月 26 日 ( 告示第 2480 号 ) 一部改正 1 定義組換え型 F11 線毛抗原産生大腸菌及びベロ細胞毒性抗原産生大腸菌の培養菌液を不活化し その遠心上清を濃縮後 油性アジュバントを添加したワクチンである
鶏大腸菌症 ( 組換え型 F11 線毛抗原 ベロ細胞毒性抗原 )( 油性アジュバント加 ) 不活化ワクチン平成 20 年 6 月 6 日 ( 告示第 913 号 ) 新規追加 平成 25 年 9 月 26 日 ( 告示第 2480 号 ) 一部改正 1 定義組換え型 F11 線毛抗原産生大腸菌及びベロ細胞毒性抗原産生大腸菌の培養菌液を不活化し その遠心上清を濃縮後 油性アジュバントを添加したワクチンである
博士学位論文審査報告書
 5 氏 名満仲翔一 学 位 の 種 類博士 ( 理学 ) 報 告 番 号甲第 465 号 学位授与年月日 2017 年 9 月 19 日 学位授与の要件学位規則 ( 昭和 28 年 4 月 1 日文部省令第 9 号 ) 第 4 条第 1 項該当 学位論文題目腸管出血性大腸菌 O157:H7 Sakai 株に存在する Stx2 ファー ジにコードされた Small Regulatory RNA SesR
5 氏 名満仲翔一 学 位 の 種 類博士 ( 理学 ) 報 告 番 号甲第 465 号 学位授与年月日 2017 年 9 月 19 日 学位授与の要件学位規則 ( 昭和 28 年 4 月 1 日文部省令第 9 号 ) 第 4 条第 1 項該当 学位論文題目腸管出血性大腸菌 O157:H7 Sakai 株に存在する Stx2 ファー ジにコードされた Small Regulatory RNA SesR
実験操作方法
 実験操作方法 A. サンプル ( 菌液 ) の調製検出目的の菌種に適した方法でサンプル調製を行い 最終的に100μl 程度の菌懸濁液を用意してください このサンプル液のうち40μlを B.EMA 処理 に使用します 残りは氷上もしくは4 で保存し EMA 処理なしサンプルを用意するために40μlを B.EMA 処理 8) 加熱殺菌 ( 不活化処理 ) の操作を行った後 C. 核酸抽出 に使用します
実験操作方法 A. サンプル ( 菌液 ) の調製検出目的の菌種に適した方法でサンプル調製を行い 最終的に100μl 程度の菌懸濁液を用意してください このサンプル液のうち40μlを B.EMA 処理 に使用します 残りは氷上もしくは4 で保存し EMA 処理なしサンプルを用意するために40μlを B.EMA 処理 8) 加熱殺菌 ( 不活化処理 ) の操作を行った後 C. 核酸抽出 に使用します
Mighty TA-cloning Kit/Mighty TA-cloning Kit for PrimeSTAR
 製品コード 6028 製品コード 6029 Mighty TA-cloning Kit ( 製品コード 6028) Mighty TA-cloning Kit for PrimeSTAR ( 製品コード 6029) 説明書 Taq DNA ポリメラーゼなどをベースとする PCR 酵素を用いて得られた増幅産物のほとんどは その 3 末端にデオキシリボアデノシン (da) が一塩基付加されています これらの
製品コード 6028 製品コード 6029 Mighty TA-cloning Kit ( 製品コード 6028) Mighty TA-cloning Kit for PrimeSTAR ( 製品コード 6029) 説明書 Taq DNA ポリメラーゼなどをベースとする PCR 酵素を用いて得られた増幅産物のほとんどは その 3 末端にデオキシリボアデノシン (da) が一塩基付加されています これらの
 ,...~,.'~ 表 2.6.2.2-26 試験管内 PAE 菌株薬剤 MIC (µg/ml) PAE (h) 1 MIC 4 MIC STFX 0.025 0.92 2.35 S. aureus FDA 209-P LVFX 0.20 0.68 2.68 CPFX 0.20 1.05 1.59 SPFX 0.10 0.35 1.07 STFX 0.025 2.33 1.14 E. coli KL-16
,...~,.'~ 表 2.6.2.2-26 試験管内 PAE 菌株薬剤 MIC (µg/ml) PAE (h) 1 MIC 4 MIC STFX 0.025 0.92 2.35 S. aureus FDA 209-P LVFX 0.20 0.68 2.68 CPFX 0.20 1.05 1.59 SPFX 0.10 0.35 1.07 STFX 0.025 2.33 1.14 E. coli KL-16
本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因
 HbA1c 測定系について ~ 原理と特徴 ~ 一般社団法人日本臨床検査薬協会 技術運営委員会副委員長 安部正義 本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因 HPLC 法 HPLC 法原理 高速液体クロマトグラフィー 混合物の分析法の一つ 固体または液体の固定相 ( 吸着剤 ) 中で 液体または気体の移動相 ( 展開剤 ) に試料を加えて移動させ
HbA1c 測定系について ~ 原理と特徴 ~ 一般社団法人日本臨床検査薬協会 技術運営委員会副委員長 安部正義 本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因 HPLC 法 HPLC 法原理 高速液体クロマトグラフィー 混合物の分析法の一つ 固体または液体の固定相 ( 吸着剤 ) 中で 液体または気体の移動相 ( 展開剤 ) に試料を加えて移動させ
発現ベクターによるRNAi
 発現ベクターによる RNAi 特長 合成 sirna を導入した場合に比べ 長期に RNAi 効果を観察できる 細胞中に高効率で sirna を発現させることが可能 研究目的に応じて任意のベクターが選択できる 原理及び説明遺伝子発現をノックダウンする手法としてRNA 干渉作用 (RNAi) が注目されている RNAi は二本鎖 RNA(dsRNA) を細胞に導入することにより 標的遺伝子のmRNAを分解し
発現ベクターによる RNAi 特長 合成 sirna を導入した場合に比べ 長期に RNAi 効果を観察できる 細胞中に高効率で sirna を発現させることが可能 研究目的に応じて任意のベクターが選択できる 原理及び説明遺伝子発現をノックダウンする手法としてRNA 干渉作用 (RNAi) が注目されている RNAi は二本鎖 RNA(dsRNA) を細胞に導入することにより 標的遺伝子のmRNAを分解し
PrimeScript® II 1st strand cDNA Synthesis Kit
 研究用 PrimeScript II 1st strand cdna Synthesis Kit 説明書 v201208da 目次 I. 概要... 3 II. キットの内容... 4 III. 保存... 4 IV. 1st-strand cdna 合成反応... 5 V. RT-PCR を行う場合... 6 VI. RNA サンプルの調製について... 6 VII. 関連製品... 7 VIII.
研究用 PrimeScript II 1st strand cdna Synthesis Kit 説明書 v201208da 目次 I. 概要... 3 II. キットの内容... 4 III. 保存... 4 IV. 1st-strand cdna 合成反応... 5 V. RT-PCR を行う場合... 6 VI. RNA サンプルの調製について... 6 VII. 関連製品... 7 VIII.
In vivo へのトライ
 アデノウイルスベクターによる RNAi 特長 高い感染能を利用し 効率よく一過性の RNAi 実験が行える 広い動物種に用いることができ 増殖細胞だけでなく静止期の細胞にも感染 発現できる 神経系を含む多くの分化 未分化動物培養細胞をターゲットにすることができ さらに動物個体への直接注入 投与による遺伝子発現が可能である 原理説明 アデノウイルスは最もよく研究されているウイルスのひとつである
アデノウイルスベクターによる RNAi 特長 高い感染能を利用し 効率よく一過性の RNAi 実験が行える 広い動物種に用いることができ 増殖細胞だけでなく静止期の細胞にも感染 発現できる 神経系を含む多くの分化 未分化動物培養細胞をターゲットにすることができ さらに動物個体への直接注入 投与による遺伝子発現が可能である 原理説明 アデノウイルスは最もよく研究されているウイルスのひとつである
1. Caov-3 細胞株 A2780 細胞株においてシスプラチン単剤 シスプラチンとトポテカン併用添加での殺細胞効果を MTS assay を用い検討した 2. Caov-3 細胞株においてシスプラチンによって誘導される Akt の活性化に対し トポテカンが影響するか否かを調べるために シスプラチ
 ( 様式甲 5) 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 朝日通雄 恒遠啓示 副査副査 瀧内比呂也谷川允彦 副査 勝岡洋治 主論文題名 Topotecan as a molecular targeting agent which blocks the Akt and VEGF cascade in platinum-resistant ovarian cancers ( 白金製剤耐性卵巣癌における
( 様式甲 5) 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 朝日通雄 恒遠啓示 副査副査 瀧内比呂也谷川允彦 副査 勝岡洋治 主論文題名 Topotecan as a molecular targeting agent which blocks the Akt and VEGF cascade in platinum-resistant ovarian cancers ( 白金製剤耐性卵巣癌における
はじめてのリアルタイムPCR
 はじめてのリアルタイム PCR 1. はじめにリアルタイム PCR 法は PCR 増幅産物の増加をリアルタイムでモニタリングし 解析する技術です エンドポイントで PCR 増幅産物を確認する従来の PCR 法に比べて 1DNA や RNA の正確な定量ができること 2 電気泳動が不要なので迅速かつ簡便に解析できコンタミネーションの危険性も小さいことなど多くの利点があります 今や 遺伝子発現解析や SNP
はじめてのリアルタイム PCR 1. はじめにリアルタイム PCR 法は PCR 増幅産物の増加をリアルタイムでモニタリングし 解析する技術です エンドポイントで PCR 増幅産物を確認する従来の PCR 法に比べて 1DNA や RNA の正確な定量ができること 2 電気泳動が不要なので迅速かつ簡便に解析できコンタミネーションの危険性も小さいことなど多くの利点があります 今や 遺伝子発現解析や SNP
図 B 細胞受容体を介した NF-κB 活性化モデル
 60 秒でわかるプレスリリース 2007 年 12 月 17 日 独立行政法人理化学研究所 免疫の要 NF-κB の活性化シグナルを増幅する機構を発見 - リン酸化酵素 IKK が正のフィーッドバックを担当 - 身体に病原菌などの異物 ( 抗原 ) が侵入すると 誰にでも備わっている免疫システムが働いて 異物を認識し 排除するために さまざまな反応を起こします その一つに 免疫細胞である B 細胞が
60 秒でわかるプレスリリース 2007 年 12 月 17 日 独立行政法人理化学研究所 免疫の要 NF-κB の活性化シグナルを増幅する機構を発見 - リン酸化酵素 IKK が正のフィーッドバックを担当 - 身体に病原菌などの異物 ( 抗原 ) が侵入すると 誰にでも備わっている免疫システムが働いて 異物を認識し 排除するために さまざまな反応を起こします その一つに 免疫細胞である B 細胞が
リアルタイムPCRの基礎知識
 1. リアルタイム PCR の用途リアルタイム PCR 法は 遺伝子発現解析の他に SNPs タイピング 遺伝子組み換え食品の検査 ウイルスや病原菌の検出 導入遺伝子のコピー数の解析などさまざまな用途に応用されている 遺伝子発現解析のような定量解析は まさにリアルタイム PCR の得意とするところであるが プラス / マイナス判定だけの定性的な解析にもその威力を発揮する これは リアルタイム PCR
1. リアルタイム PCR の用途リアルタイム PCR 法は 遺伝子発現解析の他に SNPs タイピング 遺伝子組み換え食品の検査 ウイルスや病原菌の検出 導入遺伝子のコピー数の解析などさまざまな用途に応用されている 遺伝子発現解析のような定量解析は まさにリアルタイム PCR の得意とするところであるが プラス / マイナス判定だけの定性的な解析にもその威力を発揮する これは リアルタイム PCR
Untitled
 上原記念生命科学財団研究報告集, 26 (2012) 112. 水痘帯状疱疹ウイルス感染機構の解明 定岡知彦 Key words: 水痘帯状疱疹ウイルス, 感染性ウイルス粒子産生 神戸大学大学院医学研究科感染症センター臨床ウイルス学 緒言水痘帯状疱疹ウイルス (varicella-zoster virus: VZV) は, ヒトに感染して初感染においては全身性に水痘を発症し, 脊髄後根神経節に潜伏した後,
上原記念生命科学財団研究報告集, 26 (2012) 112. 水痘帯状疱疹ウイルス感染機構の解明 定岡知彦 Key words: 水痘帯状疱疹ウイルス, 感染性ウイルス粒子産生 神戸大学大学院医学研究科感染症センター臨床ウイルス学 緒言水痘帯状疱疹ウイルス (varicella-zoster virus: VZV) は, ヒトに感染して初感染においては全身性に水痘を発症し, 脊髄後根神経節に潜伏した後,
DNA Fragmentation Kit
 研究用 DNA Fragmentation Kit 説明書 v201712da 本製品は 超音波破砕装置などの特殊な装置を使用せず 酵素処理によってゲノム DNA 等の長鎖 DNA をランダムに断片化し さらに平滑化するためのキットです 平滑化した断片はそのまま平滑末端ベクターに組み込むことができます また 平滑化を必要としない場合は 断片化までで反応を止めることもできます メチル化 DNA 濃縮や高速シーケンス解析のための前処理にも
研究用 DNA Fragmentation Kit 説明書 v201712da 本製品は 超音波破砕装置などの特殊な装置を使用せず 酵素処理によってゲノム DNA 等の長鎖 DNA をランダムに断片化し さらに平滑化するためのキットです 平滑化した断片はそのまま平滑末端ベクターに組み込むことができます また 平滑化を必要としない場合は 断片化までで反応を止めることもできます メチル化 DNA 濃縮や高速シーケンス解析のための前処理にも
Untitled
 上原記念生命科学財団研究報告集, 25 (2011) 124. 次世代タンパク質量分析と遺伝学的手法との融合によるクロマチン構造制御による DNA 修復機構の解明 廣田耕志 Key words:dna 修復, プロテオミクス,SILAC, DT40, 遺伝学 京都大学大学院医学研究科放射線遺伝学教室 緒言クロマチン構造は,DNA を規則正しく折り畳み核内にコンパクトに格納する際に重要である. 一方,
上原記念生命科学財団研究報告集, 25 (2011) 124. 次世代タンパク質量分析と遺伝学的手法との融合によるクロマチン構造制御による DNA 修復機構の解明 廣田耕志 Key words:dna 修復, プロテオミクス,SILAC, DT40, 遺伝学 京都大学大学院医学研究科放射線遺伝学教室 緒言クロマチン構造は,DNA を規則正しく折り畳み核内にコンパクトに格納する際に重要である. 一方,
研究成果報告書
 様式 C-19 科学研究費補助金研究成果報告書 平成 23 年 3 月 28 日現在 機関番号 :3714 研究種目 : 若手研究 研究期間 :28~21 課題番号 :279342 研究課題名 ( 和文 )Toll-like receptor 1 のリガンド探索および機能解析研究課題名 ( 英文 )Functional analysis of Toll-like receptor 1 研究代表者清水隆
様式 C-19 科学研究費補助金研究成果報告書 平成 23 年 3 月 28 日現在 機関番号 :3714 研究種目 : 若手研究 研究期間 :28~21 課題番号 :279342 研究課題名 ( 和文 )Toll-like receptor 1 のリガンド探索および機能解析研究課題名 ( 英文 )Functional analysis of Toll-like receptor 1 研究代表者清水隆
Bacterial 16S rDNA PCR Kit
 研究用 Bacterial 16S rdna PCR Kit 説明書 v201307da 微生物の同定は 形態的特徴 生理 生化学的性状 化学分類学的性状などを利用して行われますが これらの方法では同定までに時間を要します また 同定が困難な場合や正しい結果が得られない場合もあります 近年 微生物同定にも分子生物学を利用した方法が採用されるようになり 微生物の持つ DNA を対象として解析を行う方法が活用されています
研究用 Bacterial 16S rdna PCR Kit 説明書 v201307da 微生物の同定は 形態的特徴 生理 生化学的性状 化学分類学的性状などを利用して行われますが これらの方法では同定までに時間を要します また 同定が困難な場合や正しい結果が得られない場合もあります 近年 微生物同定にも分子生物学を利用した方法が採用されるようになり 微生物の持つ DNA を対象として解析を行う方法が活用されています
DNA Ligation Kit <Mighty Mix>
 製品コード 623 研究用 DNA Ligation Kit < Mighty Mix > 説明書 v21512da DNA フラグメントのライゲーションは 遺伝子操作実験で頻繁に行う操作です DNA 間の結合には T4 DNA Ligase が用いられますが DNA の末端構造によって反応速度は著しく異なります そのため DNA の末端形状にあわせて加える酵素量や反応時間を調節する必要があります
製品コード 623 研究用 DNA Ligation Kit < Mighty Mix > 説明書 v21512da DNA フラグメントのライゲーションは 遺伝子操作実験で頻繁に行う操作です DNA 間の結合には T4 DNA Ligase が用いられますが DNA の末端構造によって反応速度は著しく異なります そのため DNA の末端形状にあわせて加える酵素量や反応時間を調節する必要があります
SSH資料(徳島大学総合科学部 佐藤)
") だ液タンパク質 (α- アミラーゼ ) の分析 ~ 電気泳動による決定と酵素活性の測定 ~ 人の体はさまざまなタンパク質から成り立っている タンパク質とは 20 種のアミノ酸という物質が 100 個以上重合して出来る生体高分子である 髪の毛 爪 皮ふなどもタンパク質からできており 生体内のタンパク質には これ以外にもいろいろな種類と機能がある このうち ある化学反応を触媒するタンパク質を特に酵素 (
だ液タンパク質 (α- アミラーゼ ) の分析 ~ 電気泳動による決定と酵素活性の測定 ~ 人の体はさまざまなタンパク質から成り立っている タンパク質とは 20 種のアミノ酸という物質が 100 個以上重合して出来る生体高分子である 髪の毛 爪 皮ふなどもタンパク質からできており 生体内のタンパク質には これ以外にもいろいろな種類と機能がある このうち ある化学反応を触媒するタンパク質を特に酵素 (
く 細胞傷害活性の無い CD4 + ヘルパー T 細胞が必須と判明した 吉田らは 1988 年 C57BL/6 マウスが腹腔内に移植した BALB/c マウス由来の Meth A 腫瘍細胞 (CTL 耐性細胞株 ) を拒絶すること 1991 年 同種異系移植によって誘導されるマクロファージ (AIM
 を拒絶すること 1991 年 同種異系移植によって誘導されるマクロファージ (AIM") ( 様式甲 5) 氏 名 山名秀典 ( ふりがな ) ( やまなひでのり ) 学 位 の 種 類 博士 ( 医学 ) 学位授与番号 甲 第 号 学位審査年月日 平成 26 年 7 月 30 日 学位授与の要件 学位規則第 4 条第 1 項該当 Down-regulated expression of 学位論文題名 monocyte/macrophage major histocompatibility
( 様式甲 5) 氏 名 山名秀典 ( ふりがな ) ( やまなひでのり ) 学 位 の 種 類 博士 ( 医学 ) 学位授与番号 甲 第 号 学位審査年月日 平成 26 年 7 月 30 日 学位授与の要件 学位規則第 4 条第 1 項該当 Down-regulated expression of 学位論文題名 monocyte/macrophage major histocompatibility
 クルマエビホワイトスポット病の防除対策 Table 1. List of WSSV confirmed naturally and/or experimentally infected host species クルマエビホワイトスポット病の防除対策 Fig. 8. Disease control in shrimp hatchery. に 組換え大腸菌で発現した WSSV 構造タンパク質 第
クルマエビホワイトスポット病の防除対策 Table 1. List of WSSV confirmed naturally and/or experimentally infected host species クルマエビホワイトスポット病の防除対策 Fig. 8. Disease control in shrimp hatchery. に 組換え大腸菌で発現した WSSV 構造タンパク質 第
MightyAmp™ DNA Polymerase Ver.3
 研究用 MightyAmp DNA Polymerase Ver.3 説明書 v201805da MightyAmp DNA Polymerase は 究極の反応性を追求して開発された PCR 酵素であり 通常の PCR 酵素では増幅が困難な PCR 阻害物質を多く含むクルードな生体粗抽出液を用いる場合にも その強力な増幅能により良好な反応性を示します MightyAmp DNA Polymerase
研究用 MightyAmp DNA Polymerase Ver.3 説明書 v201805da MightyAmp DNA Polymerase は 究極の反応性を追求して開発された PCR 酵素であり 通常の PCR 酵素では増幅が困難な PCR 阻害物質を多く含むクルードな生体粗抽出液を用いる場合にも その強力な増幅能により良好な反応性を示します MightyAmp DNA Polymerase
16S (V3-V4) Metagenomic Library Construction Kit for NGS
 Metagenomic Library Construction Kit for NGS") 研究用 16S (V3-V4) Metagenomic Library Construction Kit for NGS 説明書 v201705da 本製品は イルミナ社 MiSeq で 16S rrna 細菌叢解析を行うための PCR 増幅キットです 細菌 16S rrna 遺伝子の V3-V4 領域を対象とし PCR 酵素に 多様な配列を効率よく増幅可能な Tks Gflex DNA Polymerase
研究用 16S (V3-V4) Metagenomic Library Construction Kit for NGS 説明書 v201705da 本製品は イルミナ社 MiSeq で 16S rrna 細菌叢解析を行うための PCR 増幅キットです 細菌 16S rrna 遺伝子の V3-V4 領域を対象とし PCR 酵素に 多様な配列を効率よく増幅可能な Tks Gflex DNA Polymerase
医薬品タンパク質は 安全性の面からヒト型が常識です ではなぜ 肌につける化粧品用コラーゲンは ヒト型でなくても良いのでしょうか? アレルギーは皮膚から 最近の学説では 皮膚から侵入したアレルゲンが 食物アレルギー アトピー性皮膚炎 喘息 アレルギー性鼻炎などのアレルギー症状を引き起こすきっかけになる
 化粧品用コラーゲンの原料 現在は 魚由来が中心 かつては ウシの皮膚由来がほとんど BSE 等病原体混入の危険 人に感染する病原体をもたない アレルギーの問題は未解決 ( むしろ問題は大きくなったかもしれない ) アレルギーを引き起こす可能性 医薬品タンパク質は 安全性の面からヒト型が常識です ではなぜ 肌につける化粧品用コラーゲンは ヒト型でなくても良いのでしょうか? アレルギーは皮膚から 最近の学説では
化粧品用コラーゲンの原料 現在は 魚由来が中心 かつては ウシの皮膚由来がほとんど BSE 等病原体混入の危険 人に感染する病原体をもたない アレルギーの問題は未解決 ( むしろ問題は大きくなったかもしれない ) アレルギーを引き起こす可能性 医薬品タンパク質は 安全性の面からヒト型が常識です ではなぜ 肌につける化粧品用コラーゲンは ヒト型でなくても良いのでしょうか? アレルギーは皮膚から 最近の学説では
1_alignment.ppt
 " " " " n " n n " n " n n n " n n n n " n LGPSSKQTGKGW-SRIWDN! + +! LN-ITKSAGKGAIMRLGDA! " n -------TGKG--------!! -------AGKG--------! " n w w w " n w w " " " 11 12 " n w w w " n w w A! M! O! A!
" " " " n " n n " n " n n n " n n n n " n LGPSSKQTGKGW-SRIWDN! + +! LN-ITKSAGKGAIMRLGDA! " n -------TGKG--------!! -------AGKG--------! " n w w w " n w w " " " 11 12 " n w w w " n w w A! M! O! A!
論文の内容の要旨
 1. 2. 3. 4. 5. 6. WASP-interacting protein(wip) CR16 7. 8..pdf Adobe Acrobat WINDOWS2000 論文の内容の要旨 論文題目 WASP-interacting protein(wip) ファミリー遺伝子 CR16 の機能解析 氏名坂西義史 序 WASP(Wiskott-Aldrich syndrome protein)
1. 2. 3. 4. 5. 6. WASP-interacting protein(wip) CR16 7. 8..pdf Adobe Acrobat WINDOWS2000 論文の内容の要旨 論文題目 WASP-interacting protein(wip) ファミリー遺伝子 CR16 の機能解析 氏名坂西義史 序 WASP(Wiskott-Aldrich syndrome protein)
食糧 その科学と技術 No.45(2007.03)
") PCR γ Bacillus subtilis natto Bacillus subtilis Bacillus subtilis Bacillus subtilis 168 γ Insertion sequence γ L D α γ Bacillus Bacillus subtilis Bacillus licheniformis Bacillus anthracis Bacillus megaterium
PCR γ Bacillus subtilis natto Bacillus subtilis Bacillus subtilis Bacillus subtilis 168 γ Insertion sequence γ L D α γ Bacillus Bacillus subtilis Bacillus licheniformis Bacillus anthracis Bacillus megaterium
Microsoft Word - Gateway technology_J1.doc
 テクノロジー Gateway の基本原理 テクノロジーは λ ファージが大腸菌染色体へ侵入する際に関与する部位特異的組換えシステムを基礎としています (Ptashne, 1992) テクノロジーでは λ ファージの組換えシステムのコンポーネントを改変することで 組み換え反応の特異性および効率を高めています (Bushman et al, 1985) このセクションでは テクノロジーの基礎となっている
テクノロジー Gateway の基本原理 テクノロジーは λ ファージが大腸菌染色体へ侵入する際に関与する部位特異的組換えシステムを基礎としています (Ptashne, 1992) テクノロジーでは λ ファージの組換えシステムのコンポーネントを改変することで 組み換え反応の特異性および効率を高めています (Bushman et al, 1985) このセクションでは テクノロジーの基礎となっている
PowerPoint プレゼンテーション
 酵素 : タンパク質の触媒 タンパク質 Protein 酵素 Enzyme 触媒 Catalyst 触媒 Cataylst: 特定の化学反応の反応速度を速める物質 自身は反応の前後で変化しない 酵素 Enzyme: タンパク質の触媒 触媒作用を持つタンパク質 第 3 回 : タンパク質はアミノ酸からなるポリペプチドである 第 4 回 : タンパク質は様々な立体構造を持つ 第 5 回 : タンパク質の立体構造と酵素活性の関係
酵素 : タンパク質の触媒 タンパク質 Protein 酵素 Enzyme 触媒 Catalyst 触媒 Cataylst: 特定の化学反応の反応速度を速める物質 自身は反応の前後で変化しない 酵素 Enzyme: タンパク質の触媒 触媒作用を持つタンパク質 第 3 回 : タンパク質はアミノ酸からなるポリペプチドである 第 4 回 : タンパク質は様々な立体構造を持つ 第 5 回 : タンパク質の立体構造と酵素活性の関係
untitled
 ORYZA POLYAMINE ORYZA POLYAMINE -P -PC -LC 1. Ver.1.0 MM ver. 1.0MM 1) Roseeuw D.I. et al., Epidermal keratinocytes actively maintain their intracellular polyamine levels. Cell Tissue Kinet. 16, 493-504
ORYZA POLYAMINE ORYZA POLYAMINE -P -PC -LC 1. Ver.1.0 MM ver. 1.0MM 1) Roseeuw D.I. et al., Epidermal keratinocytes actively maintain their intracellular polyamine levels. Cell Tissue Kinet. 16, 493-504
Microsoft PowerPoint - 資料6-1_高橋委員(公開用修正).pptx
.pptx") 第 1 回遺伝子治療等臨床研究に関する指針の見直しに関する専門委員会 平成 29 年 4 月 12 日 ( 水 ) 資料 6-1 ゲノム編集技術の概要と問題点 筑波大学生命科学動物資源センター筑波大学医学医療系解剖学発生学研究室 WPI-IIIS 筑波大学国際睡眠医科学研究機構筑波大学生命領域学際研究 (TARA) センター 高橋智 ゲノム編集技術の概要と問題点 ゲノム編集とは? なぜゲノム編集は遺伝子改変に有効?
第 1 回遺伝子治療等臨床研究に関する指針の見直しに関する専門委員会 平成 29 年 4 月 12 日 ( 水 ) 資料 6-1 ゲノム編集技術の概要と問題点 筑波大学生命科学動物資源センター筑波大学医学医療系解剖学発生学研究室 WPI-IIIS 筑波大学国際睡眠医科学研究機構筑波大学生命領域学際研究 (TARA) センター 高橋智 ゲノム編集技術の概要と問題点 ゲノム編集とは? なぜゲノム編集は遺伝子改変に有効?
糖鎖の新しい機能を発見:補体系をコントロールして健康な脳神経を維持する
 糖鎖の新しい機能を発見 : 補体系をコントロールして健康な脳神経を維持する ポイント 神経細胞上の糖脂質の糖鎖構造が正常パターンになっていないと 細胞膜の構造や機能が障害されて 外界からのシグナルに対する反応や攻撃に対する防御反応が異常になることが示された 細胞膜のタンパク質や脂質に結合している糖鎖の役割として 補体の活性のコントロールという新規の重要な機能が明らかになった 糖脂質の糖鎖が欠損すると
糖鎖の新しい機能を発見 : 補体系をコントロールして健康な脳神経を維持する ポイント 神経細胞上の糖脂質の糖鎖構造が正常パターンになっていないと 細胞膜の構造や機能が障害されて 外界からのシグナルに対する反応や攻撃に対する防御反応が異常になることが示された 細胞膜のタンパク質や脂質に結合している糖鎖の役割として 補体の活性のコントロールという新規の重要な機能が明らかになった 糖脂質の糖鎖が欠損すると
Pyrobest ® DNA Polymerase
 Pyrobest DNA Polymerase 説明書 v201102da 対活Pyrobest DNA Polymerase は Pyrococcus sp. 由来の 3' 5' exonuclease 活性 (proof reading 活性 ) を有する耐熱性 α 型 DNA ポリメラーゼです α 型 DNA ポリメラーゼは Pol I 型ポリメラーゼ (Taq DNA ポリメラーゼなど )
Pyrobest DNA Polymerase 説明書 v201102da 対活Pyrobest DNA Polymerase は Pyrococcus sp. 由来の 3' 5' exonuclease 活性 (proof reading 活性 ) を有する耐熱性 α 型 DNA ポリメラーゼです α 型 DNA ポリメラーゼは Pol I 型ポリメラーゼ (Taq DNA ポリメラーゼなど )
Untitled
 上原記念生命科学財団研究報告集, 26 (2012) 3. 糸状菌の未利用遺伝子発現による天然物創出 大島吉輝 Key words: 糸状菌, 二次代謝物, 未利用遺伝子, エピジェネティクス 東北大学大学院薬学研究科医薬資源化学分野 緒言医薬品開発にとって重要な役割を果たしている糸状菌 ( カビ類 ) のゲノム解読が進むにつれ, 二次代謝物の生産に関わる多くの未利用生合成遺伝子の存在が明らかになってきた.
上原記念生命科学財団研究報告集, 26 (2012) 3. 糸状菌の未利用遺伝子発現による天然物創出 大島吉輝 Key words: 糸状菌, 二次代謝物, 未利用遺伝子, エピジェネティクス 東北大学大学院薬学研究科医薬資源化学分野 緒言医薬品開発にとって重要な役割を果たしている糸状菌 ( カビ類 ) のゲノム解読が進むにつれ, 二次代謝物の生産に関わる多くの未利用生合成遺伝子の存在が明らかになってきた.
 Sulfonylurea Receptors Pancreatic Type: SUR1 Cardiac Type: SUR2A Smooth Muscle Type: SUR2B Kir Channels uka-rp-1/kir6.1 BIR/Kir6.2 1) Ho K, et al : Cloning and expression of an inwardly rectifying ATP-regulated
Sulfonylurea Receptors Pancreatic Type: SUR1 Cardiac Type: SUR2A Smooth Muscle Type: SUR2B Kir Channels uka-rp-1/kir6.1 BIR/Kir6.2 1) Ho K, et al : Cloning and expression of an inwardly rectifying ATP-regulated
遺伝子検査の基礎知識
 リアルタイム PCR( インターカレーター法 ) 実験ガイドこの文書では インターカレーター法 (TB Green 検出 ) によるリアルタイム PCR について 蛍光検出の原理や実験操作の流れなどを解説します 実際の実験操作の詳細については 各製品の取扱説明書をご参照ください - 目次 - 1 蛍光検出の原理 2 実験に必要なもの 3 実験操作法 4 結果の解析 1 1 蛍光検出の原理 インターカレーターによる蛍光検出の原理
リアルタイム PCR( インターカレーター法 ) 実験ガイドこの文書では インターカレーター法 (TB Green 検出 ) によるリアルタイム PCR について 蛍光検出の原理や実験操作の流れなどを解説します 実際の実験操作の詳細については 各製品の取扱説明書をご参照ください - 目次 - 1 蛍光検出の原理 2 実験に必要なもの 3 実験操作法 4 結果の解析 1 1 蛍光検出の原理 インターカレーターによる蛍光検出の原理
( 様式甲 5) 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 教授 森脇真一 井上善博 副査副査 教授教授 東 治 人 上 田 晃 一 副査 教授 朝日通雄 主論文題名 Transgene number-dependent, gene expression rate-independe
 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 教授 森脇真一 井上善博 副査副査 教授教授 東 治 人 上 田 晃 一 副査 教授 朝日通雄 主論文題名 Transgene number-dependent, gene expression rate-independe") ( 様式甲 5) 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 森脇真一 井上善博 副査副査 東 治 人 上 田 晃 一 副査 朝日通雄 主論文題名 Transgene number-dependent, gene expression rate-independent rejection of D d -, K d -, or D d K d -transgened mouse skin
( 様式甲 5) 学位論文内容の要旨 論文提出者氏名 論文審査担当者 主査 森脇真一 井上善博 副査副査 東 治 人 上 田 晃 一 副査 朝日通雄 主論文題名 Transgene number-dependent, gene expression rate-independent rejection of D d -, K d -, or D d K d -transgened mouse skin
Microsoft PowerPoint - 4_河邊先生_改.ppt
 組換え酵素を用いた配列部位 特異的逐次遺伝子導入方法 Accumulative gene integration system using recombinase 工学研究院化学工学部門河邉佳典 2009 年 2 月 27 日 < 研究背景 > 1 染色体上での遺伝子増幅の有用性 動物細胞での場合 新鮮培地 空気 + 炭酸ガス 使用済み培地 医薬品タンパク質を生産する遺伝子を導入 目的遺伝子の多重化
組換え酵素を用いた配列部位 特異的逐次遺伝子導入方法 Accumulative gene integration system using recombinase 工学研究院化学工学部門河邉佳典 2009 年 2 月 27 日 < 研究背景 > 1 染色体上での遺伝子増幅の有用性 動物細胞での場合 新鮮培地 空気 + 炭酸ガス 使用済み培地 医薬品タンパク質を生産する遺伝子を導入 目的遺伝子の多重化
Bacterial 16S rDNA Clone Library Construction Kit
 研究用 Bacterial 16S rdna Clone Library Construction Kit 説明書 v201303da_2 Bacterial 16S rdna Clone Library Construction Kit は 環境中のバクテリアの群集構造を解析する手法の 1 つである 16S rdna Clone Library 法を効率良く行うために開発された タカラバイオ独自のシステムです
研究用 Bacterial 16S rdna Clone Library Construction Kit 説明書 v201303da_2 Bacterial 16S rdna Clone Library Construction Kit は 環境中のバクテリアの群集構造を解析する手法の 1 つである 16S rdna Clone Library 法を効率良く行うために開発された タカラバイオ独自のシステムです
3'-Full RACE Core Set
 研究用 3'-Full RACE Core Set 説明書 v201703da RNA の解析において RT-PCR を利用することにより目的の領域を増幅した後 クローニングやシーケンスを行うことが可能です しかしながら mrna から完全長の cdna を得ることは困難であることが多く この様な場合 得られた cdna の情報を元にさらに上流や下流をクローニングする RACE(Rapid Amplification
研究用 3'-Full RACE Core Set 説明書 v201703da RNA の解析において RT-PCR を利用することにより目的の領域を増幅した後 クローニングやシーケンスを行うことが可能です しかしながら mrna から完全長の cdna を得ることは困難であることが多く この様な場合 得られた cdna の情報を元にさらに上流や下流をクローニングする RACE(Rapid Amplification
Taro-04-1(雌雄判別)
") 高嶋康晴 Yasuharu TAKASHIMA 要 約 多くの PCR 法では 電気泳動により分離された特定塩基長の PCR 産物や制限酵素で断片化された DNA 断片の塩基長のパターンを目視で検出し生物種や品種等の判定を行う 電気泳動にかかる一連の操作を手作業で行うことによる分析者への負担の軽減及び分析処理の効率化を図るために マグロ属魚類及びサケ科魚類の魚種判別法について アガロースゲル電気泳動と全自動電気泳動装置による電気泳動の比較検討を行った
高嶋康晴 Yasuharu TAKASHIMA 要 約 多くの PCR 法では 電気泳動により分離された特定塩基長の PCR 産物や制限酵素で断片化された DNA 断片の塩基長のパターンを目視で検出し生物種や品種等の判定を行う 電気泳動にかかる一連の操作を手作業で行うことによる分析者への負担の軽減及び分析処理の効率化を図るために マグロ属魚類及びサケ科魚類の魚種判別法について アガロースゲル電気泳動と全自動電気泳動装置による電気泳動の比較検討を行った
Untitled
 上原記念生命科学財団研究報告集, 22(2008) 113. アテローム動脈硬化に関与する単球 / マクロファージ特異的 microrna 群の同定 深尾太郎 Key words: 動脈硬化,microRNA, マクロファージ, 炎症, マウス * 東京大学大学院工学系研究科化学生命工学専攻 緒言動脈硬化は, 先進国における死因群の主たる原因となる重大な疾患である. 加齢や生活習慣がその発症や程度に大きく関わることから,
上原記念生命科学財団研究報告集, 22(2008) 113. アテローム動脈硬化に関与する単球 / マクロファージ特異的 microrna 群の同定 深尾太郎 Key words: 動脈硬化,microRNA, マクロファージ, 炎症, マウス * 東京大学大学院工学系研究科化学生命工学専攻 緒言動脈硬化は, 先進国における死因群の主たる原因となる重大な疾患である. 加齢や生活習慣がその発症や程度に大きく関わることから,
Microsoft PowerPoint - プレゼンテーション1
 A A RNA からタンパク質へ mrna の塩基配列は 遺伝暗号を介してタンパク質のアミノ酸の配列へと翻訳される trna とアミノ酸の結合 RNA 分子は 3 通りの読み枠で翻訳できる trnaは アミノ酸とコドンを結びつけるアダプター分子である (Ψ; プソイドウリジン D; ジヒドロウリジンどちらもウラシルが化学修飾したもの ) アミノアシル trna 合成酵素によって アミノ酸と trna
A A RNA からタンパク質へ mrna の塩基配列は 遺伝暗号を介してタンパク質のアミノ酸の配列へと翻訳される trna とアミノ酸の結合 RNA 分子は 3 通りの読み枠で翻訳できる trnaは アミノ酸とコドンを結びつけるアダプター分子である (Ψ; プソイドウリジン D; ジヒドロウリジンどちらもウラシルが化学修飾したもの ) アミノアシル trna 合成酵素によって アミノ酸と trna
< 背景 > HMGB1 は 真核生物に存在する分子量 30 kda の非ヒストン DNA 結合タンパク質であり クロマチン構造変換因子として機能し 転写制御および DNA の修復に関与します 一方 HMGB1 は 組織の損傷や壊死によって細胞外へ分泌された場合 炎症性サイトカイン遺伝子の発現を増強
 岡山大学記者クラブ文部科学記者会科学記者会 御中 平成 30 年 3 月 22 日岡山大学 歯周炎進行のメカニズムの一端を解明 歯周炎による骨吸収が抗 HMGB1 抗体投与により抑制 岡山大学大学院医歯薬学総合研究科の平田千暁医員 ( 当時 ) 山城圭介助教 高柴正悟教授 ( 以上 歯周病態学分野 ) と西堀正洋教授 ( 薬理学分野 ) の研究グループは 歯周炎の進行に炎症メディエーター 1 である
岡山大学記者クラブ文部科学記者会科学記者会 御中 平成 30 年 3 月 22 日岡山大学 歯周炎進行のメカニズムの一端を解明 歯周炎による骨吸収が抗 HMGB1 抗体投与により抑制 岡山大学大学院医歯薬学総合研究科の平田千暁医員 ( 当時 ) 山城圭介助教 高柴正悟教授 ( 以上 歯周病態学分野 ) と西堀正洋教授 ( 薬理学分野 ) の研究グループは 歯周炎の進行に炎症メディエーター 1 である
Dr, Fujita
 免疫組織化学染色における抗原賦活の原理について考える 東京医科大学病理学講座 藤田浩司 はじめにパラフィン切片により証明される抗原の種類は, この十数年のうちに飛躍的に増加した. 元来, 免疫組織化学法は凍結切片やアセトンを用いた固定により抗原温存を図っていたが, 現在ではホルマリン固定材料のパラフィン切片によっても多くの抗原の同定が可能となった. その理由には, 1) ホルマリン固定に抵抗性の抗原を認識するモノクローナル抗体の開発
免疫組織化学染色における抗原賦活の原理について考える 東京医科大学病理学講座 藤田浩司 はじめにパラフィン切片により証明される抗原の種類は, この十数年のうちに飛躍的に増加した. 元来, 免疫組織化学法は凍結切片やアセトンを用いた固定により抗原温存を図っていたが, 現在ではホルマリン固定材料のパラフィン切片によっても多くの抗原の同定が可能となった. その理由には, 1) ホルマリン固定に抵抗性の抗原を認識するモノクローナル抗体の開発
るが AML 細胞における Notch シグナルの正確な役割はまだわかっていない mtor シグナル伝達系も白血病細胞の増殖に関与しており Palomero らのグループが Notch と mtor のクロストークについて報告している その報告によると 活性型 Notch が HES1 の発現を誘導
 学位論文の内容の要旨 論文提出者氏名 奥橋佑基 論文審査担当者 主査三浦修副査水谷修紀 清水重臣 論文題目 NOTCH knockdown affects the proliferation and mtor signaling of leukemia cells ( 論文内容の要旨 ) < 要旨 > 目的 : sirna を用いた NOTCH1 と NOTCH2 の遺伝子発現の抑制の 白血病細胞の細胞増殖と下流のシグナル伝達系に対する効果を解析した
学位論文の内容の要旨 論文提出者氏名 奥橋佑基 論文審査担当者 主査三浦修副査水谷修紀 清水重臣 論文題目 NOTCH knockdown affects the proliferation and mtor signaling of leukemia cells ( 論文内容の要旨 ) < 要旨 > 目的 : sirna を用いた NOTCH1 と NOTCH2 の遺伝子発現の抑制の 白血病細胞の細胞増殖と下流のシグナル伝達系に対する効果を解析した
パナテスト ラットβ2マイクログロブリン
 研究用試薬 2014 年 4 月作成 EIA 法ラット β 2 マイクログロブリン測定キット PRH111 パナテスト A シリーズラット β 2- マイクロク ロフ リン 1. はじめに β 2 - マイクログロブリンは, 血液, 尿, および体液中に存在し, ヒトでは腎糸球体障害, 自己免疫疾患, 悪性腫瘍, 肝疾患などによって血中濃度が変化するといわれています. また,β 2 - マイクログロブリンの尿中濃度は,
研究用試薬 2014 年 4 月作成 EIA 法ラット β 2 マイクログロブリン測定キット PRH111 パナテスト A シリーズラット β 2- マイクロク ロフ リン 1. はじめに β 2 - マイクログロブリンは, 血液, 尿, および体液中に存在し, ヒトでは腎糸球体障害, 自己免疫疾患, 悪性腫瘍, 肝疾患などによって血中濃度が変化するといわれています. また,β 2 - マイクログロブリンの尿中濃度は,
Taro-kv20025.jtd
 猫ウイルス性鼻気管炎 猫カリシウイルス感染症 2 価 猫汎白血球減少症 猫白血病 ( 猫白血病ウイルス由来防御抗原たん白遺伝子導入カナリア痘ウイルス ) 猫クラミジア感染症混合ワクチン 平成 24 年 7 月 4 日 ( 告示第 1623 号 ) 新規追加 弱毒猫ウイルス性鼻気管炎ウイルス 弱毒猫汎白血球減少症ウイルス及び弱毒猫クラミドフィラ フェリスをそれぞれ培養細胞で増殖させて得たウイルス液と2
猫ウイルス性鼻気管炎 猫カリシウイルス感染症 2 価 猫汎白血球減少症 猫白血病 ( 猫白血病ウイルス由来防御抗原たん白遺伝子導入カナリア痘ウイルス ) 猫クラミジア感染症混合ワクチン 平成 24 年 7 月 4 日 ( 告示第 1623 号 ) 新規追加 弱毒猫ウイルス性鼻気管炎ウイルス 弱毒猫汎白血球減少症ウイルス及び弱毒猫クラミドフィラ フェリスをそれぞれ培養細胞で増殖させて得たウイルス液と2
Powered by TCPDF ( Title ハイリスクHPV 型のタンパクを標的とした新たな分子標的治療に関する基礎的検討 Sub Title Study of a new therapy targeting proteins of high-risk HPV Au
 Powered by TCPDF (www.tcpdf.org) Title ハイリスクHPV 型のタンパクを標的とした新たな分子標的治療に関する基礎的検討 Sub Title Study of a new therapy targeting proteins of high-risk HPV Author 大野, 暁子 (Ono, Akiko) Publisher Publication year
Powered by TCPDF (www.tcpdf.org) Title ハイリスクHPV 型のタンパクを標的とした新たな分子標的治療に関する基礎的検討 Sub Title Study of a new therapy targeting proteins of high-risk HPV Author 大野, 暁子 (Ono, Akiko) Publisher Publication year
Multiplex PCR Assay Kit Ver. 2
 研究用 Multiplex PCR Assay Kit Ver. 2 説明書 v201510da マルチプレックス PCR は 一つの PCR 反応系に複数のプライマー対を同時に使用することで 複数の遺伝子領域を同時に増幅する方法です マルチプレックス PCR を行うことで 試薬や機材の節約による経済性 同時検出による迅速性でのメリットに加え 貴重なサンプルの有効利用も可能です 本キットは高速にプライミングする酵素とプライマーのアニーリングの特異性を極限まで高めた反応液組成とを組み合わせたマルチプレックス
研究用 Multiplex PCR Assay Kit Ver. 2 説明書 v201510da マルチプレックス PCR は 一つの PCR 反応系に複数のプライマー対を同時に使用することで 複数の遺伝子領域を同時に増幅する方法です マルチプレックス PCR を行うことで 試薬や機材の節約による経済性 同時検出による迅速性でのメリットに加え 貴重なサンプルの有効利用も可能です 本キットは高速にプライミングする酵素とプライマーのアニーリングの特異性を極限まで高めた反応液組成とを組み合わせたマルチプレックス
<4D F736F F D B82C982C282A282C482512E646F63>
 サンプル条件および固定化分子の選択 Biacoreの実験ではセンサーチップに固定化する分子をリガンド それに対して結合を測定する分子をアナライトと呼びます いずれの分子をリガンドとし アナライトとするかは 実験系を構築する上で重要です 以下にサンプルに適したリガンド アナライトの設計方法やサンプルの必要条件などをご紹介します アナライト リガンド センサーチップ (1) タンパク質リガンドとしてもアナライトとしても用いることができます
サンプル条件および固定化分子の選択 Biacoreの実験ではセンサーチップに固定化する分子をリガンド それに対して結合を測定する分子をアナライトと呼びます いずれの分子をリガンドとし アナライトとするかは 実験系を構築する上で重要です 以下にサンプルに適したリガンド アナライトの設計方法やサンプルの必要条件などをご紹介します アナライト リガンド センサーチップ (1) タンパク質リガンドとしてもアナライトとしても用いることができます
報道発表資料 2006 年 4 月 13 日 独立行政法人理化学研究所 抗ウイルス免疫発動機構の解明 - 免疫 アレルギー制御のための新たな標的分子を発見 - ポイント 異物センサー TLR のシグナル伝達機構を解析 インターフェロン産生に必須な分子 IKK アルファ を発見 免疫 アレルギーの有効
 60 秒でわかるプレスリリース 2006 年 4 月 13 日 独立行政法人理化学研究所 抗ウイルス免疫発動機構の解明 - 免疫 アレルギー制御のための新たな標的分子を発見 - がんやウイルスなど身体を蝕む病原体から身を守る物質として インターフェロン が注目されています このインターフェロンのことは ご存知の方も多いと思いますが 私たちが生まれながらに持っている免疫をつかさどる物質です 免疫細胞の情報の交換やウイルス感染に強い防御を示す役割を担っています
60 秒でわかるプレスリリース 2006 年 4 月 13 日 独立行政法人理化学研究所 抗ウイルス免疫発動機構の解明 - 免疫 アレルギー制御のための新たな標的分子を発見 - がんやウイルスなど身体を蝕む病原体から身を守る物質として インターフェロン が注目されています このインターフェロンのことは ご存知の方も多いと思いますが 私たちが生まれながらに持っている免疫をつかさどる物質です 免疫細胞の情報の交換やウイルス感染に強い防御を示す役割を担っています
Microsoft Word - 04H22研究報告岡久.doc
 報文 和菓子の賞味期限予測のための耐熱性芽胞菌増殖予測モデルの開発 Development of Bacterial Spore Growth Predictive Model for Predictive at Best-Before Date of Japanese-Style Confection 岡久修己 * Naoki Okahisa 抄録市販の和菓子等から分離した耐熱性芽胞菌 42 株から,
報文 和菓子の賞味期限予測のための耐熱性芽胞菌増殖予測モデルの開発 Development of Bacterial Spore Growth Predictive Model for Predictive at Best-Before Date of Japanese-Style Confection 岡久修己 * Naoki Okahisa 抄録市販の和菓子等から分離した耐熱性芽胞菌 42 株から,