Microsoft PowerPoint - aaseq_09Apr28 [互換モード]
|
|
|
- なおちか まるこ
- 6 years ago
- Views:
Transcription
1 近畿大学 農学部 生命情報学 蛋白質の物理化学的性質とアミノ酸配列解析 2009 年 4 月 28 日 ( 火 ) 奈良先端大 情報 蛋白質機能予測学講座川端猛 takawaba@is.naist.jp 平成 21 年度 生命情報学 & 生命情報学実習 講義日程 講義生命情報学演習生命情報学演習 4/7 川端 1 配列決定とバイオインフォマティクス 4/14 川端 2 ペアワイズアライメントと配列相同性検索 川端 主要 WEBデータベースの使用法 (BLAST) 4/21 川端 3 マルチプルアライメントとその応用中村 hemffice を用いた計算化学演習 4/28 川端 4 蛋白質の物理化学的性質と配列解 析 5/12 川端 5 分子系統学基礎 中村 系統樹作成演習 (lustalx) /19 川端 6 蛋白質立体構造データの情報解析川端蛋白質立体構造データの可視化 (asmol) 5/26 川端 7 >> 試験 << 6/2 金谷 1 ポストゲノム解析入門 ( トランスクリプトーム解析 ) 6/9 金谷 2 ポストゲノム解析入門 ( インタラクトーム解 金谷 1 発現プロフィール解析演習 析 ) 6/16 金谷 3 ポストゲノム解析 ( 統合解析 ) 金谷 2 インタラクトローム 代謝物解析演習 6/23 金谷 4 メタボローム解析 ( その1) 6/30 金谷 5 メタボローム解析 ( その2) 7/7 金谷 6 メタボローム解析 ( その3) 7/14 金谷 7 >> 試験 << 1


2 配列比較によるタンパク質の分類 Pfam : 蛋白質ファミリのデータベース 各蛋白質ファミリのマルチプルアライメント MM などを集めたデータベース 2
3 配列比較によるタンパク質分類 1970 年代から 配列の一致度 (sequence identity) に基づいて グループを作る作業が行われてきた どのくらいの類似性で一つにまとめるかは様々な考えがあるが 少なくとも 相同なタンパク質群 を一つのグループにまとめるべき ファミリー (family, 族 ): : 進化的に関係のある ( 相同な ) タンパク質グループのこと 複数のファミリーがあつまった大きなグループ : スーパーファミリー (superfamily, clan) 一つのファミリーの中の小さなグループ : サブファミリー (subfamily) family subfamily superfamily 配列 1 配列 2 A 1 ドメイン単位による分類 配列の部分どうしだけが類似していた場合 配列 3 A 2 B 2 B 3 3 ドメインに分割すると 部分一致を重視すると すべて一つのグループになるが A を共有 配列 1 配列 2 配列 3 B を共有 1 と 2 が類似 2 と 3 が類似していても 1 と 3 は類似していない! A 1 A 1 B 2 3 A 2 B 2 A 2 B 3 B 3 3 きれいに分類することができる!! 3
4 Pkinase_Tyr ドメインをもつタンパク質の例 Family : Pkinase_Tyr (PF07714) : Protein tyrosine kinase K_UMA ABL_UMA BTK_UMA ILK_UMA METK_UMA PGFB_UMA Pfam データベース ( からの引用 蛋白質の物理化学的性質とアミノ酸配列解析 4
 様々な物理化学的な性質を持った側鎖が主鎖のペプチド結合で連なる Leu(L) (")
 Q 負荷電 E K 正荷電 親水性 ys() ( 親水性 )")
 という現象 温度や変性剤濃度によって可逆に変化 変性状態")


5 蛋白質 : ペプチド結合したアミノ酸群 Phe(F) ( 疎水性 ) Ala(A) ( 疎水性 ) 様々な物理化学的な性質を持った側鎖が主鎖のペプチド結合で連なる Leu(L) ( 疎水性 ) A P 脂肪族 G I V L Y M F W 疎水性 芳香族 S T D Thr ( 親水性 ) Q 負荷電 E K 正荷電 親水性 ys() ( 親水性 ) アミノ酸は全部で 20 種類 Val(V) ( 疎水性 ) Asp(D) ( 親水性 ) フォールディング ( 折り畳み ) という現象 温度や変性剤濃度によって可逆に変化 変性状態 (D) 非常に多種の構造の集合大きく広がっている 天然状態 () ほとんど唯一の構造小さくコンパクトに折りたたまっている 折り畳みは 原則としてその蛋白質以外の分子の介助を必要としない アミノ酸配列の情報だけで 天然状態の立体構造が決定される どうやってコンパクトになるか? (1) 主鎖の水素結合 (2) 側鎖間の疎水性相互作用 あるアミノ酸配列がどうやって一つの構造を決めるのか??? 5
6 アミノ酸の疎水性の分布による蛋白質の特徴づけ タンパク質の構造 性質による大きな分類 水溶性タンパク質 soluble protein 膜タンパク質 membrane protein 球状タンパク質 globular protein 不定形タンパク質 Intrinsically disordered protein タンパク質の例 多くの酵素 抗体 転写因子など 真核生物の転写因子の非ドメイン領域など 配列上の特徴 疎水性 親水性のアミノ酸がバランスよく含まれている 親水性アミノ酸が多い 特に EPQSK が多い レセプター トラン疎水性アミノ酸が多く スポーターなど連続する疎水性アミノ酸領域 ( 膜貫通へリックス ) が観察されることが多い 6


")



7 球状タンパク質の疎水性アミノ酸 球状タンパク質では 分子内部に疎水性アミノ酸 分子表面に親水性アミノ酸が分布する傾向にある 疎水基 :AILMFWV 親水基 :DEQGKPSTY 5p21:rasp21 7pcyA:plastocyanin 1mbdA:Myoglobin 1timA:Triose phosphate isomerase 疎水性相互作用 (hydrophobic interaction) : 水分子と親和性の少ない非極性 (non-polar 疎水性) 基が水溶液中で互いに集まろうとする相互作用 電気や磁気と異なり符号はなく 疎水性どうしのものは相手を問わず集合する 球状タンパク質の疎水性アミノ酸 球状タンパク質では 分子内部に疎水性アミノ酸 分子表面に親水性アミノ酸が分布する傾向にある 疎水基 :AILMFWV 親水基 :DEQGKPSTY 1mbdA:Myoglobin 7pcyA:plastocyanin 1timA:Triose phosphate isomerase 疎水性相互作用 (hydrophobic interaction) : 水分子と親和性の少ない非極性 (non-polar 疎水性 ) 基が水溶液中で互いに集まろうとする相互作用 電気や磁気と異なり符号はなく 疎水性どうしのものは相手を問わず集合する 7

![5 方法 配列を横軸にとって 縦軸に対応する疎水性指標をプロット 前後数残基でスムージングジング w 1 V [ i] = KD{ Seq[ i + w]} 2w](/docs-images/71/66222707/images/8-2.jpg "+ 1 k = w http://kr.expasy.org/tools/protscale.")
8 疎水性指標による内外予測 Kyte and Doolittle(1982) の疎水性指標 I 4.5 V 4.2 L 3.8 F M 1.9 A 1.8 G -0.4 T -0.7 S -0.8 W -0.9 Y -1.3 P Q E -3.5 D -3.5 K 方法 配列を横軸にとって 縦軸に対応する疎水性指標をプロット 前後数残基でスムージングジング w 1 V [ i] = KD{ Seq[ i + w]} 2w + 1 k = w 予測結果と実際の埋もれ度との比較 1mbd:Myoglobin 8

9 膜タンパク質 脂質頭部 ( 親水性 ) 膜タンパク質 : 脂質二重膜に埋没して機能するタンパク質 膜タンパク質 脂質尾部 ( 疎水性 ) 性質 (1) 疎水的な脂質尾部と相互作用するため タンパク質表面には疎水性アミノ酸が多い (2) 水に溶けにくく 凝集しやすい (3) 多くは膜貫通へリックスを持つ (4) 膜貫通へリックスの長さは アミノ酸ぐらい アミノ酸配列からも連続した疎水性アミノ酸領域としてある程度 予測可能 不定形タンパク質 intrinsically disordered protein 親水性アミノ酸が多く 疎水性アミノ酸が少ないため 天然状態でもコンパクトに折たたまらない 親水性アミノ酸の中でも EPQSK 球状タンパク質ドメインをつなぐ領域としてよく見られる 真核生物に多く 原核生物には少ない 転写因子に特に多くみられる 生物学的な機能はよくわかっていないが タンパク質間相互作用に重要な働きをするといわれている 9
10 タンパク質の構造 性質による大きな分類 水溶性タンパク質 soluble protein 膜タンパク質 membrane protein 球状タンパク質 globular protein 不定形タンパク質 Intrinsically disordered protein タンパク質の例 多くの酵素 抗体 転写因子など 真核生物の転写因子の非ドメイン領域など 配列上の特徴 疎水性 親水性のアミノ酸がバランスよく含まれている 親水性アミノ酸が多い 特に EPQSK が多い レセプター トラン疎水性アミノ酸が多く スポーターなど連続する疎水性アミノ酸領域 ( 膜貫通へリックス ) が観察されることが多い アミノ酸疎水性による配列特徴解析 : 学籍番号 : 氏名 : (1) 以下の配列の疎水性アミノ酸 (AFILMVW) を で囲み そのタンパク質のタイプを判定せよ (2) へリックスの周期的疎水パターン X X X,, を で囲め >1mbdA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q >1bm1A ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) P E W I W L A L G T A L M G L G T L Y F L V K G M G V S D P D A K K F Y A I T T L V P A I A F T M Y L S M L L G Y G L T M V P F G G E Q P I Y W A Y A D W L F T T P L L L L D L A L L V D A D Q G T I L A L V G A D G I M I G T G L V G A L T K V Y S Y F V W W A I S T A A M L Y I L Y V L F F G F T S K A E S M P E V A S T F K V L V T V V L W S A Y P V V W L I G S E G A G I V P L I E T L L F M V L D V S A K V G F G L I L L S >Z428_UMA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) M T E T E P A E T G G Y A S L E E D D E D L S P G P E S S D S E Y T L S E P D S E E E E D E E E E E E E T T D D P E Y D P G Y K V K Q L G G G G G P S A P A A Q P P A Q P Q L G S P L G E A P P G T P P L P A T A P Q E A P A P E G A L G E E E E E P P A G E G P A G E E E E E E E E E G T Y T E E D S F D L G E L G F M L A G E V 10
11 アミノ酸疎水性による配列特徴解析 : 学籍番号 : 氏名 : (1) 以下の配列の疎水性アミノ酸 (AFILMVW) を で囲み そのタンパク質のタイプを判定せよ (2) へリックスの周期的疎水パターン X X X,, を で囲め >1mbdA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q >1bm1A ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) P E W I W L A L G T A L M G L G T L Y F L V K G M G V S D P D A K K F Y A I T T L V P A I A F T M Y L S M L L G Y G L T M V P F G G E Q P I Y W A Y A D W L F T T P L L L L D L A L L V D A D Q G T I L A L V G A D G I M I G T G L V G A L T K V Y S Y F V W W A I S T A A M L Y I L Y V L F F G F T S K A E S M P E V A S T F K V L V T V V L W S A Y P V V W L I G S E G A G I V P L I E T L L F M V L D V S A K V G F G L I L L S >Z428_UMA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) M T E T E P A E T G G Y A S L E E D D E D L S P G P E S S D S E Y T L S E P D S E E E E D E E E E E E E T T D D P E Y D P G Y K V K Q L G G G G G P S A P A A Q P P A Q P Q L G S P L G E A P P G T P P L P A T A P Q E A P A P E G A L G E E E E E P P A G E G P A G E E E E E E E E E G T Y T E E D S F D L G E L G F M L A G E V アミノ酸疎水性による配列特徴解析 : 学籍番号 : 氏名 : (1) 以下の配列の疎水性アミノ酸 (AFILMVW) を で囲み そのタンパク質のタイプを判定せよ (2) へリックスの周期的疎水パターン X X X,, を で囲め >1mbdA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q >1bm1A ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) P E W I W L A L G T A L M G L G T L Y F L V K G M G V S D P D A K K F Y A I T T L V P A I A F T M Y L S M L L G Y G L T M V P F G G E Q P I Y W A Y A D W L F T T P L L L L D L A L L V D A D Q G T I L A L V G A D G I M I G T G L V G A L T K V Y S Y F V W W A I S T A A M L Y I L Y V L F F G F T S K A E S M P E V A S T F K V L V T V V L W S A Y P V V W L I G S E G A G I V P L I E T L L F M V L D V S A K V G F G L I L L S >Z428_UMA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) M T E T E P A E T G G Y A S L E E D D E D L S P G P E S S D S E Y T L S E P D S E E E E D E E E E E E E T T D D P E Y D P G Y K V K Q L G G G G G P S A P A A Q P P A Q P Q L G S P L G E A P P G T P P L P A T A P Q E A P A P E G A L G E E E E E P P A G E G P A G E E E E E E E E E G T Y T E E D S F D L G E L G F M L A G E V 11
( どれか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K")






12 アミノ酸疎水性による配列特徴解析 : 学籍番号 : 氏名 : (1) 以下の配列の疎水性アミノ酸 (AFILMVW) を で囲み そのタンパク質のタイプを判定せよ (2) へリックスの周期的疎水パターン X X X,, を で囲め >1mbdA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q >1bm1A ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) P E W I W L A L G T A L M G L G T L Y F L V K G M G V S D P D A K K F Y A I T T L V P A I A F T M Y L S M L L G Y G L T M V P F G G E Q P I Y W A Y A D W L F T T P L L L L D L A L L V D A D Q G T I L A L V G A D G I M I G T G L V G A L T K V Y S Y F V W W A I S T A A M L Y I L Y V L F F G F T S K A E S M P E V A S T F K V L V T V V L W S A Y P V V W L I G S E G A G I V P L I E T L L F M V L D V S A K V G F G L I L L S >Z428_UMA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) M T E T E P A E T G G Y A S L E E D D E D L S P G P E S S D S E Y T L S E P D S E E E E D E E E E E E E T T D D P E Y D P G Y K V K Q L G G G G G P S A P A A Q P P A Q P Q L G S P L G E A P P G T P P L P A T A P Q E A P A P E G A L G E E E E E P P A G E G P A G E E E E E E E E E G T Y T E E D S F D L G E L G F M L A G E V アミノ酸配列から膜貫通へリックスを予測するサーバ SSUI サーバ 12
13 蛋白質の二次構造 (α へリックス β シート ) 二次構造 : 主鎖の規則的な水素結合 主鎖は 基と 基の繰り返しであるため 極めて親水的 13
14 二次構造 : 主鎖の規則的な水素結合 主鎖は 基と 基の繰り返しであるため 極めて親水的 水分子が水素結合しやすい 二次構造 : 主鎖の規則的な水素結合 主鎖は 基と 基の繰り返しであるため 極めて親水的 水分子が水素結合しやすい 折りたたまれ 水が排除されると水素結合は切断される エネルギー的に不利 14



15 二次構造 : 主鎖の規則的な水素結合 主鎖は 基と 基の繰り返しであるため 極めて親水的 水分子が水素結合しやすい 折りたたまれ 水が排除されると水素結合は切断される エネルギー的に不利 蛋白質内で新たな水素結合を作る必要性 二次構造の生成 主鎖の水素結合 :α へリックス α i+4 α i+3 α i-1 i 番目のがi+4 番目の と水素結合を形成 α 3.6 残基が 1 周期の右巻きらせん =が同じ向きに並ぶため へリックス全体に電気双極子が発生 i ー + 末 末 15




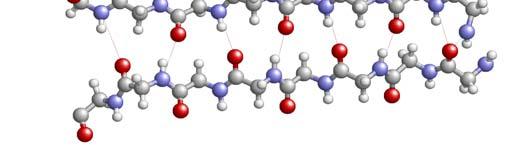

16 主鎖の水素結合 : 逆平行 β シート β ヘアピン βターン β ターン β ヘアピン 主鎖の水素結合 : 平行 β シート β α β モチーフ 平行 β シートが α へリックスで接続された構造モチーフ 16





17 1mbd 球状タンパク質の構造クラス all-α α/β 1n55A 1pqwA α- へリックスが中心 α- へリックスと β- シートが配列上交互に現れる 平行 β- シートが中心 1mqk all-β 1fxd α+β 1a2p β- シートが中心逆平行が多い α- へリックスと β- シートが混在 配列上の並びの規則はない逆平行と平行の β- シートが混在 アミノ酸ごとに 2 次構造のなりやすさに差がある α ヘリックス hou-fasmann のパラメータ P Glu (scop %list:3077chains を用いて再計算した値 ) ( α) = f ( Glu α) f ( α) A 1.42 E 1.37 L 1.33 Q 1.30 M K W 1.07 I 1.06 F 0.97 Y 0.96 V D T 0.77 S G 0.43 P 0.42 β シート コイル V 1.95 I 1.77 F 1.46 Y W 1.29 T 1.20 L 1.11 M S 0.83 K 0.79 Q 0.77 A 0.76 E 0.70 G D 0.53 P 0.42 P 1.66 G D 1.33 S T K Q 0.90 E 0.88 Y 0.84 W 0.83 F 0.82 M 0.81 A 0.81 L 0.72 V 0.65 I
18 (3) α ヘリックスを好むアミノ酸 AEQLM を で囲みなさい この二つが all-α か all-β のどちらかだとしたら どちらに属すると考えられるか をつけて答えよ >1mbdA (all-αクラス all-βクラス )( どちらか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA (all-αクラス all-βクラス ) ( どちらか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q all-α 生命情報学 ( ) all-β (3) α ヘリックスを好むアミノ酸 AEQLM を で囲みなさい この二つが all-α か all-β のどちらかだとしたら どちらに属すると考えられるか をつけて答えよ >1mbdA (all-αクラス all-βクラス )( どちらか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA (all-αクラス all-βクラス ) ( どちらか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q all-α 生命情報学 ( ) all-β 18
( どちらか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q")

 A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A")

 K A F L M I L Q Y 両親媒性ヘリックス : 片側が疎水的")
19 (3) α ヘリックスを好むアミノ酸 AEQLM を で囲みなさい この二つが all-α か all-β のどちらかだとしたら どちらに属すると考えられるか をつけて答えよ >1mbdA (all-αクラス all-βクラス )( どちらか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA (all-αクラス all-βクラス ) ( どちらか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q all-α 生命情報学 ( ) all-β 疎水性の車輪図 (elical Wheel) 1mbdA の最後のヘリックスの配列 :ADAQGAMKALELFKDIAAKYKEL E D A L A D K K K G E A A A Myoglobin (1mbdA) K A F L M I L Q Y 両親媒性ヘリックス : 片側が疎水的 反対の側が親水的になっているヘリックスのこと へリックスの 1 回転の周期が 3.6 アミノ酸なので 3 から 4 アミノ酸周期の疎水性 ヘリックス構造を示唆 19
 以下の配列の疎水性アミノ酸 (AFILMVW) を で囲み そのタンパク質のタイプを判定せよ (2) へリックスの周期的疎水パターン X X X,, を で囲め >1mbdA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) V L S E G E W Q L V L V W A K")
( どれか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y")

20 2 次構造ごとに疎水性パターンに特徴がある 疎水 親水 Kawabata, T. and Doi, J.(1997) "Improvement of Protein Secondary Structure Prediction Using Binary Word Encoding", Proteins, Vol 27, pp アミノ酸疎水性による配列特徴解析 : 学籍番号 : 氏名 : (1) 以下の配列の疎水性アミノ酸 (AFILMVW) を で囲み そのタンパク質のタイプを判定せよ (2) へリックスの周期的疎水パターン X X X,, を で囲め >1mbdA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q >1bm1A ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) P E W I W L A L G T A L M G L G T L Y F L V K G M G V S D P D A K K F Y A I T T L V P A I A F T M Y L S M L L G Y G L T M V P F G G E Q P I Y W A Y A D W L F T T P L L L L D L A L L V D A D Q G T I L A L V G A D G I M I G T G L V G A L T K V Y S Y F V W W A I S T A A M L Y I L Y V L F F G F T S K A E S M P E V A S T F K V L V T V V L W S A Y P V V W L I G S E G A G I V P L I E T L L F M V L D V S A K V G F G L I L L S >Z428_UMA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) M T E T E P A E T G G Y A S L E E D D E D L S P G P E S S D S E Y T L S E P D S E E E E D E E E E E E E T T D D P E Y D P G Y K V K Q L G G G G G P S A P A A Q P P A Q P Q L G S P L G E A P P G T P P L P A T A P Q E A P A P E G A L G E E E E E P P A G E G P A G E E E E E E E E E G T Y T E E D S F D L G E L G F M L A G E V 20

21 アミノ酸疎水性による配列特徴解析 : 学籍番号 : 氏名 : (1) 以下の配列の疎水性アミノ酸 (AFILMVW) を で囲み そのタンパク質のタイプを判定せよ (2) へリックスの周期的疎水パターン X X X,, を で囲め >1mbdA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) V L S E G E W Q L V L V W A K V E A D V A G G Q D I L I L F K S P E T L E K F D F K L K T E A E M K A S E D L K K G V T V L T A L G A I L K K K G E A E L K P L A Q S A T K K I P I K Y L E F I S E A I I V L S P G D F G A D A Q G A M K A L E L F K D I A A K Y K E L G Y Q G >7pcyA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) A A I V K L G G D D G S L A F V P I T V G A G E S I E F I A G F P I V F D E D A V P A G V D A D A I S A E D Y L S K G Q T V V K L T T P G T Y G V Y D P S G A G M K M T I T V Q >1bm1A ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) P E W I W L A L G T A L M G L G T L Y F L V K G M G V S D P D A K K F Y A I T T L V P A I A F T M Y L S M L L G Y G L T M V P F G G E Q P I Y W A Y A D W L F T T P L L L L D L A L L V D A D Q G T I L A L V G A D G I M I G T G L V G A L T K V Y S Y F V W W A I S T A A M L Y I L Y V L F F G F T S K A E S M P E V A S T F K V L V T V V L W S A Y P V V W L I G S E G A G I V P L I E T L L F M V L D V S A K V G F G L I L L S >Z428_UMA ( 球状タンパク質 膜タンパク質 不定形タンパク質 )( どれか一つを選択 ) M T E T E P A E T G G Y A S L E E D D E D L S P G P E S S D S E Y T L S E P D S E E E E D E E E E E E E T T D D P E Y D P G Y K V K Q L G G G G G P S A P A A Q P P A Q P Q L G S P L G E A P P G T P P L P A T A P Q E A P A P E G A L G E E E E E P P A G E G P A G E E E E E E E E E G T Y T E E D S F D L G E L G F M L A G E V 2 次構造予測 アミノ酸配列から 2 次構造を予測すること アミノ酸配列 2 次構造 TEVVGAPSIYLDGAQKLD ceeeeecccccc E β シート c コイル α ヘリックス 21
22 2 次構造予測の現状のまとめ hou-fasmann 等 70 年代に開発された方法の多くは正答率は60% 以下 ニューラルネットワークや改良 G 法を用いると 63 ~68% ぐらいの正答率 マルチプルアライメントを入力とする方法は正答率は70% を超える 現状では76% ぐらいまで到達 一般にαへリックスに比べ βシートの予測は困難 現状のベストの方法マルチプルアライメントを入力とするニューラルネットワーク Blast,lustalW, PSI-BLAST,MMer アンサンブル学習再帰ネットワーク K- との組み合わせ 2 次構造予測の例 Single : 1 つの配列を入力とするニューラルネットワーク [Q3=66.6%] Profile : 複数の配列を入力とするニューラルネットワーク [Q3=83.5%] >1n55A [c.1.1.1] ISMEASE A: TISEPSPATE ISMEASE AAseq :AKPQPIAAAWKGTTASIEKLVQVFETISDVQVVAPTFVIPLVQAKLPKYV Single :ccccccccccccccccccccccceeeeeccccccccccccccc Profile:ccccEEEEEccccccccccccccEEEEEcccccccE bserve:ccccceeeeeccccccccccccceeeeeccccccccccee AAseq :ISAQAIAKSGAFTGEVSMPILKDIGVWVILGSETYYGETDEIVAQKVSEAKQGF Single :ccccccccccccccccccccccceeeeeccccccccccccccc Profile:EEccccccccccccccccccccEEEEccccccccccccccc bserve:eeecccccccccccccccccceeeeccccccccc AAseq :MVIAIGETLQQEAQTAKVVLSQTSAIAAKLTKDAWQVVLAYEPVWAIGTGKVATPE Single :EEEEEccccEEccccccccEEcccccccc Profile:cEEEEEcccccccccccccccccEEEEEccccccccccccc bserve:eeeeeecccccccccccccccceeeeeccccccccccccc AAseq :QAQEVLLLKWVSEIGTDVAAKLILYGGSVAAAATLYAKPDIGFLVGGASLKPE Single :cccccccccccccccccccccccccccc Profile:ccccccccEEEEcccccccccccccccccccc bserve:ccceeeeeccccccccccccceeeecccccccc AAseq :FDIIDAT Si l 22


23 タンパク質の大きさと疎水性の関係 親水層の幅は一定 (6A ぐらい ) 疎水的環境 親水的環境 表面の親水層の幅はアミノ酸一つ分 (6A ぐらい) タンパク質が球形なら アミノ酸数が多くなるほど 疎水性のアミノ酸の比率が高くなってしまう 疎水性のアミノ酸が多すぎると 折り畳まる前に凝集して沈殿してしまう危険性 ドメイン構造と疎水性の関係 Tyrosine protein kinase K (1ad5A, K_UMA) S3 domain Tyrosine-kinase domain 3 つの分断された疎水性コア ドメイン構造 S2 domain 大きなタンパク質は いくつかの構造上のまとまり ( ドメイン ) を持つことが多い 23
24 参考図書 構造生物学一般について Bluce Alberis 他著 ( 中村桂子 松原謙一監訳 ) Essential 細胞生物学原書第 2 版 (2005) 第 2 章 第 4 章 南江堂 松澤洋 ( 編集 ) タンパク質工学の基礎 (2004) 東京化学同人.Branden & J.Tooze ( 勝部幸輝ら訳 ) タンパク質の構造入門 (2000), ニュートンプレス G.A.Petsko & D.inge ( 横山茂之監訳 ) タンパク質の構造と機能ゲノム時代のアプローチ (2005) メディカル サイエンス インターナショナル Arthur M.Lesk ( 高木淳一訳 ) ポストゲノム時代のタンパク質科学 構造 機能 ゲノミクスー (2007) 化学同人 構造バイオインフォマティクス 藤博幸編 はじめてのバイオインフォマティクス (2006) 講談社サイエンティフィク 郷通子 高橋健一編 基礎と実習バイオインフォマティクス (2004) 共立出版 Arthur M.Lesk( 岡崎康司 坊農秀雄監訳 ) バイオインフォマティクス基礎講義一歩進んだ発想をみがくために (2003), メディカル サイエンス インターナショナル 美宅成樹 榊佳之 バイオインフォマティクス (2003) 第 6 7 章 東京化学同人 日本バイオインフォマティクス学会編 バイオインフォマティクス事典 第 10 章 共立出版 (2006) 配列の分類 モチーフ プロフィール法 美宅成樹 榊佳之 バイオインフォマティクス (2003) 第 3,4 章 東京化学同人 日本バイオインフォマティクス学会編 バイオインフォマティクス事典 第 7 8 章 共立出版 (2006) 24
近畿大学 農学部 生命情報学 生体高分子の立体構造とその予測 Ras MTEYKLVVVGAGGVGKSAL TIQLIQNHFVDEYDPTIED SYRKQVVIDGETCLLDILD TAGQEEYSAMRDQYMRTGE GFLCVFAINNTKSFEDIHQ YREQIKRVKDSDDVP
 近畿大学 農学部 生命情報学 生体高分子の立体構造とその予測 Ras MTEYKLVVVGAGGVGKSAL TIQLIQHFVDEYDPTIED SYRKQVVIDGETCLLDILD TAGQEEYSAMRDQYMRTGE GFLCVFAITKSFEDIHQ YREQIKRVKDSDDVPMVLV GKCDLAARTVESRQAQDL ARSYGIPYIETSAKTRQGV EDAFYTLVREIRQH
近畿大学 農学部 生命情報学 生体高分子の立体構造とその予測 Ras MTEYKLVVVGAGGVGKSAL TIQLIQHFVDEYDPTIED SYRKQVVIDGETCLLDILD TAGQEEYSAMRDQYMRTGE GFLCVFAITKSFEDIHQ YREQIKRVKDSDDVPMVLV GKCDLAARTVESRQAQDL ARSYGIPYIETSAKTRQGV EDAFYTLVREIRQH
bioinfo ppt
 IT バイオインフォマティクス第 8 回 BIO 藤博幸 Outline (1) 立体構造解析の背景 (2) 立体構造データベース PDB (3) 可視化ツール (4) 構造比較と構造分類 (5) 立体構造予測 (6) 立体構造を用いた分子機能解析 構造を比較することの意味 アミノ酸配列に比べ 立体構造は保存しやすい アミノ酸配列は類似していない つまり配列レベル の検索で検出できないような類似性
IT バイオインフォマティクス第 8 回 BIO 藤博幸 Outline (1) 立体構造解析の背景 (2) 立体構造データベース PDB (3) 可視化ツール (4) 構造比較と構造分類 (5) 立体構造予測 (6) 立体構造を用いた分子機能解析 構造を比較することの意味 アミノ酸配列に比べ 立体構造は保存しやすい アミノ酸配列は類似していない つまり配列レベル の検索で検出できないような類似性
第2章 生物有機化学実験及び実験法
 1.9.2. アミノ酸配列の解析 1.9.2.1. ドメイン構造の検索 - 応用生命科学科のホームページ (http://www.biochemistry.kais.kyoto-u.ac.jp/) に入って以下の実習を行なう - 蛋白質は通常一つのドメインからなるのではなく, 多くのドメインが集まったモジュール構造をとっている. 各ドメインはドメイン単位で機能を持つと共に, 蛋白質内の他のドメイン構造と連係し,
1.9.2. アミノ酸配列の解析 1.9.2.1. ドメイン構造の検索 - 応用生命科学科のホームページ (http://www.biochemistry.kais.kyoto-u.ac.jp/) に入って以下の実習を行なう - 蛋白質は通常一つのドメインからなるのではなく, 多くのドメインが集まったモジュール構造をとっている. 各ドメインはドメイン単位で機能を持つと共に, 蛋白質内の他のドメイン構造と連係し,
生物物理 Vol. 45 No. 1 (2005) だけ正確なアラインメントが必要な方 (4) 立体構造とアミノ酸配列の関係, あるいは立体構造と機能との関係に興味がある方 2. おもなサービス 2.1 ペアワイズ3Dアラインメントこれは2つの構造をアラインメントする基本的な機能であり,MATRAS
 だけ正確なアラインメントが必要な方 (4) 立体構造とアミノ酸配列の関係, あるいは立体構造と機能との関係に興味がある方 2. おもなサービス 2.1 ペアワイズ3Dアラインメントこれは2つの構造をアラインメントする基本的な機能であり,MATRAS") 生物物理 45(1),41-44(2005) 立体構造比較サーバ MATRAS の使い方 1. はじめに 奈良先端科学技術大学院大学情報科学研究科川端猛 あるタンパク質と似ている他のタンパク質を探したいとき, まず最初に試みるべきはアミノ酸配列の相同性検索であろう. しかし, 同じファミリーのタンパク質の中でも, アミノ酸配列の一致度が低くなってくると, 配列の類似性だけで議論するのには限界が出てくる.
生物物理 45(1),41-44(2005) 立体構造比較サーバ MATRAS の使い方 1. はじめに 奈良先端科学技術大学院大学情報科学研究科川端猛 あるタンパク質と似ている他のタンパク質を探したいとき, まず最初に試みるべきはアミノ酸配列の相同性検索であろう. しかし, 同じファミリーのタンパク質の中でも, アミノ酸配列の一致度が低くなってくると, 配列の類似性だけで議論するのには限界が出てくる.
タンパク質の合成と 構造 機能 7 章 +24 頁 転写と翻訳リボソーム遺伝子の調節タンパク質の構造弱い結合とタンパク質の機能
 タンパク質の合成と 構造 機能 7 章 +24 頁 転写と翻訳リボソーム遺伝子の調節タンパク質の構造弱い結合とタンパク質の機能 タンパク質の合成 セントラル ドグマによると 遺伝子が持つ情報は タンパク質を合成することで発現 (Expression) される それは 2 段階の反応で進行する DNA 転写 (Transcription) DNA の塩基配列から mrna の塩基配列へ染色体の
タンパク質の合成と 構造 機能 7 章 +24 頁 転写と翻訳リボソーム遺伝子の調節タンパク質の構造弱い結合とタンパク質の機能 タンパク質の合成 セントラル ドグマによると 遺伝子が持つ情報は タンパク質を合成することで発現 (Expression) される それは 2 段階の反応で進行する DNA 転写 (Transcription) DNA の塩基配列から mrna の塩基配列へ染色体の
セリン OH 基は極性をもつ 親水的である トレオニン OH 基は極性をもつ 親水的である チロシン OH 基は極性をもつ 親水的である 解離してマイナスの電荷を帯びる 4 側鎖 アラニン 疎水的である グリシンの次に単純 グリシン もっとも単純な構造のアミノ酸 α 炭素が不斉炭素でないので唯一立体
 生物化学概論 Ⅰ シケプリ セントラルドグマ DNA は情報を担っている分子ではありますが それ自体は何の機能も持ちません 情報は RNA に写し取られ rrna やtRNA のように RNA として機能するか あるいは mrna として情報が写し取られ リボソームがこの情報をもとにタンパク質を合成して はじめて機能を持った分子が作られます 情報の流れは常に DNA RNA タンパク質と流れていき
生物化学概論 Ⅰ シケプリ セントラルドグマ DNA は情報を担っている分子ではありますが それ自体は何の機能も持ちません 情報は RNA に写し取られ rrna やtRNA のように RNA として機能するか あるいは mrna として情報が写し取られ リボソームがこの情報をもとにタンパク質を合成して はじめて機能を持った分子が作られます 情報の流れは常に DNA RNA タンパク質と流れていき
スライド 1
 タンパク質 ( 生化学 1) 平成 29 年 4 月 20 日病態生化学分野 分子酵素化学分野教授 山縣和也 生化学 1のスケジュール 4 月 20 日 講義開始 6 月 1 日 中間試験 9 月 25 日 生化学 1 試験 講義日程 内容は一部変更があります 講義資料 ( 山縣 吉澤分 ): 熊本大学病態生化学 で検索 ID: Biochem2 パスワード :76TgFD3Xc 生化学 1 の合否判定は
タンパク質 ( 生化学 1) 平成 29 年 4 月 20 日病態生化学分野 分子酵素化学分野教授 山縣和也 生化学 1のスケジュール 4 月 20 日 講義開始 6 月 1 日 中間試験 9 月 25 日 生化学 1 試験 講義日程 内容は一部変更があります 講義資料 ( 山縣 吉澤分 ): 熊本大学病態生化学 で検索 ID: Biochem2 パスワード :76TgFD3Xc 生化学 1 の合否判定は
Microsoft PowerPoint - protein1.ppt [互換モード]
![Microsoft PowerPoint - protein1.ppt [互換モード]](/thumbs/92/108226396.jpg "Microsoft PowerPoint - protein1.ppt [互換モード]") 生物物理化学 1 セントラルドグマ (central doguma) DA から蛋白質が作られるまでの道筋 フランシス クリックが提唱した 原核生物と真核生物では 若干関与するタンパク質が異なるが 基本的には同じメカニズムで転写 翻訳 タンパク合成が行われる 原核生物 : 核膜が無い ( 構造的に区別出来る核を持たない ) 細胞 ( これを原核細胞という ) から成る生物で 細菌類や藍藻類がこれに属する
生物物理化学 1 セントラルドグマ (central doguma) DA から蛋白質が作られるまでの道筋 フランシス クリックが提唱した 原核生物と真核生物では 若干関与するタンパク質が異なるが 基本的には同じメカニズムで転写 翻訳 タンパク合成が行われる 原核生物 : 核膜が無い ( 構造的に区別出来る核を持たない ) 細胞 ( これを原核細胞という ) から成る生物で 細菌類や藍藻類がこれに属する
第2章 生物有機化学実験及び実験法
 1.9.2.2. タンパク質立体構造解析ツールの利用 - 応用生命科学科のホームページ (http://www.biochemistry.kais.kyoto-u.ac.jp/) に入って以下の実習を行なう - タンパク質の機能を理解するためにはタンパク質の立体構造を正しく理解する必要がある. タンパク質の立体構造は主に X 線結晶構造解析により決定されている. 決定された構造情報はタンパク質データバンク
1.9.2.2. タンパク質立体構造解析ツールの利用 - 応用生命科学科のホームページ (http://www.biochemistry.kais.kyoto-u.ac.jp/) に入って以下の実習を行なう - タンパク質の機能を理解するためにはタンパク質の立体構造を正しく理解する必要がある. タンパク質の立体構造は主に X 線結晶構造解析により決定されている. 決定された構造情報はタンパク質データバンク
核内受容体遺伝子の分子生物学
 核内受容体遺伝子の分子生物学 佐賀大学農学部 助教授和田康彦 本講義のねらい 核内受容体を例として脊椎動物における分子生物学的な思考方法を体得する 核内受容体遺伝子を例として脊椎動物における遺伝子解析手法を概観する 脊椎動物における核内受容体遺伝子の役割について理解する ヒトや家畜における核内受容体遺伝子研究の応用について理解する セントラルドグマ ゲノム DNA から相補的な m RNA( メッセンシ
核内受容体遺伝子の分子生物学 佐賀大学農学部 助教授和田康彦 本講義のねらい 核内受容体を例として脊椎動物における分子生物学的な思考方法を体得する 核内受容体遺伝子を例として脊椎動物における遺伝子解析手法を概観する 脊椎動物における核内受容体遺伝子の役割について理解する ヒトや家畜における核内受容体遺伝子研究の応用について理解する セントラルドグマ ゲノム DNA から相補的な m RNA( メッセンシ
KASEAA 52(1) (2014)
 (2014)") 解説 TRP チャネルがさまざまな 刺激に応答できる仕組み * 1 * 2 * * 48 1 1 1 1 49 図 2 哺乳類 TRP チャネルの構造 情報 A TRPA1 の電子顕微鏡単粒子解析 像, 図中破線部分は構造から予測した 細 胞 膜 の 界 面 の 位 置 を 示 す B TRPV1 のアンキリンリピート C TRPM7 の α-キナーゼドメイン D TRPM7 のコイルドコイルドメイン
解説 TRP チャネルがさまざまな 刺激に応答できる仕組み * 1 * 2 * * 48 1 1 1 1 49 図 2 哺乳類 TRP チャネルの構造 情報 A TRPA1 の電子顕微鏡単粒子解析 像, 図中破線部分は構造から予測した 細 胞 膜 の 界 面 の 位 置 を 示 す B TRPV1 のアンキリンリピート C TRPM7 の α-キナーゼドメイン D TRPM7 のコイルドコイルドメイン
生命情報学
 生命情報学 (6) タンパク質構造解析 阿久津達也 京都大学化学研究所 バイオインフォマティクスセンター タンパク質立体構造 アミノ酸とタンパク質 アミノ酸 :20 種類 アミノ酸 R 側鎖 タンパク質 : アミノ酸の鎖 ( 短いものはペプチドと呼ばれる ) 蛋白質 アミノ基 H R C N H H N H C O C H H N C O OH カルボシキル基 O H C C R ペプチド結合 タンパク質の種類と高次構造
生命情報学 (6) タンパク質構造解析 阿久津達也 京都大学化学研究所 バイオインフォマティクスセンター タンパク質立体構造 アミノ酸とタンパク質 アミノ酸 :20 種類 アミノ酸 R 側鎖 タンパク質 : アミノ酸の鎖 ( 短いものはペプチドと呼ばれる ) 蛋白質 アミノ基 H R C N H H N H C O C H H N C O OH カルボシキル基 O H C C R ペプチド結合 タンパク質の種類と高次構造
<4D F736F F F696E74202D2097A791CC8D5C91A282A982E782CC8FEE95F1928A8F6F2E >
 平成 23 年 5 月 11 日構造バイオインフォマティクス基礎 立体構造からの情報抽出 東京大学大学院農学生命科学研究科アグリバイオインフォマティクス教育研究ユニット寺田透 立体構造データの使い方 個別のデータの利用 変異体の設計 立体構造予測 次回の講義で解説 複数のデータの比較 類似性に基づく機能予測 立体構造比較 データを網羅的に利用 配列と立体構造の関係 傾向を明らかにする 配列と立体構造の関係
平成 23 年 5 月 11 日構造バイオインフォマティクス基礎 立体構造からの情報抽出 東京大学大学院農学生命科学研究科アグリバイオインフォマティクス教育研究ユニット寺田透 立体構造データの使い方 個別のデータの利用 変異体の設計 立体構造予測 次回の講義で解説 複数のデータの比較 類似性に基づく機能予測 立体構造比較 データを網羅的に利用 配列と立体構造の関係 傾向を明らかにする 配列と立体構造の関係
Microsoft Word 生体分子構造学.docx
 生体分子構造学 構造から生体分子の機能の解明へ 北里大学理学部 米田茂隆 2015 夏学期 http://www.kitasato-u.ac.jp/sci/resea/buturi/seitai/yoneda/text.html アミノ酸の化学構造アミノ酸はどれも左図のような基本的な構造をもっている 左図で側鎖と書いた球がアミノ酸の種類により異なる 1 側鎖以外の部分を主鎖という 主鎖の原子は N
生体分子構造学 構造から生体分子の機能の解明へ 北里大学理学部 米田茂隆 2015 夏学期 http://www.kitasato-u.ac.jp/sci/resea/buturi/seitai/yoneda/text.html アミノ酸の化学構造アミノ酸はどれも左図のような基本的な構造をもっている 左図で側鎖と書いた球がアミノ酸の種類により異なる 1 側鎖以外の部分を主鎖という 主鎖の原子は N
生物学入門
 第 2 章生命の化学的基礎 生物を構成している元素は 地球上の物質を構成している元素と何ら異なることはない 化学で学んできたこと これから学ぶことが 生物学を理解するための基礎となる 化学の詳しいことは 化学科のおこなう講義や実習に任せることにして ここでは生物学を学ぶために必要な 最低限のことを学ぶことにする 1. 水の性質 生体を構成している分子の中で 割合が一番多いのは 水 である 生体に占める水の割合はおよそ
第 2 章生命の化学的基礎 生物を構成している元素は 地球上の物質を構成している元素と何ら異なることはない 化学で学んできたこと これから学ぶことが 生物学を理解するための基礎となる 化学の詳しいことは 化学科のおこなう講義や実習に任せることにして ここでは生物学を学ぶために必要な 最低限のことを学ぶことにする 1. 水の性質 生体を構成している分子の中で 割合が一番多いのは 水 である 生体に占める水の割合はおよそ
Dr, Fujita
 免疫組織化学染色における抗原賦活の原理について考える 東京医科大学病理学講座 藤田浩司 はじめにパラフィン切片により証明される抗原の種類は, この十数年のうちに飛躍的に増加した. 元来, 免疫組織化学法は凍結切片やアセトンを用いた固定により抗原温存を図っていたが, 現在ではホルマリン固定材料のパラフィン切片によっても多くの抗原の同定が可能となった. その理由には, 1) ホルマリン固定に抵抗性の抗原を認識するモノクローナル抗体の開発
免疫組織化学染色における抗原賦活の原理について考える 東京医科大学病理学講座 藤田浩司 はじめにパラフィン切片により証明される抗原の種類は, この十数年のうちに飛躍的に増加した. 元来, 免疫組織化学法は凍結切片やアセトンを用いた固定により抗原温存を図っていたが, 現在ではホルマリン固定材料のパラフィン切片によっても多くの抗原の同定が可能となった. その理由には, 1) ホルマリン固定に抵抗性の抗原を認識するモノクローナル抗体の開発
国際塩基配列データベース n DNA のデータベース GenBank ( アメリカ :Na,onal Center for Biotechnology Informa,on, NCBI が運営 ) EMBL ( ヨーロッパ : 欧州生命情報学研究所が運営 ) DDBJ ( 日本 : 国立遺伝研内の日
 EMBL ( ヨーロッパ : 欧州生命情報学研究所が運営 ) DDBJ ( 日本 : 国立遺伝研内の日") 生物情報工学 BioInforma*cs 3 遺伝子データベース 16/06/09 1 国際塩基配列データベース n DNA のデータベース GenBank ( アメリカ :Na,onal Center for Biotechnology Informa,on, NCBI が運営 ) EMBL ( ヨーロッパ : 欧州生命情報学研究所が運営 ) DDBJ ( 日本 : 国立遺伝研内の日本 DNA データバンクが運営
生物情報工学 BioInforma*cs 3 遺伝子データベース 16/06/09 1 国際塩基配列データベース n DNA のデータベース GenBank ( アメリカ :Na,onal Center for Biotechnology Informa,on, NCBI が運営 ) EMBL ( ヨーロッパ : 欧州生命情報学研究所が運営 ) DDBJ ( 日本 : 国立遺伝研内の日本 DNA データバンクが運営
<4D F736F F D E E8EBF B493FC96E52E646F6378>
 生体分子科学 ( 改訂 4c 版 ) 補足タンパク質ドメイン超入門 1. タンパク質構造とドメインタンパク質は 主鎖の相互作用による α-ヘリックス β-シートの形成と側鎖間相互作用によってフォールディング folding することで全体構造が決まる 完成したタンパク質構造中にはドメイン domain を認めることができる タンパク質でいうドメインとは 他のタンパク質内にも共通して存在する普遍的な機能
生体分子科学 ( 改訂 4c 版 ) 補足タンパク質ドメイン超入門 1. タンパク質構造とドメインタンパク質は 主鎖の相互作用による α-ヘリックス β-シートの形成と側鎖間相互作用によってフォールディング folding することで全体構造が決まる 完成したタンパク質構造中にはドメイン domain を認めることができる タンパク質でいうドメインとは 他のタンパク質内にも共通して存在する普遍的な機能
Microsoft PowerPoint - BIセンターセミナー2013.pptx[読み取り専用]
![Microsoft PowerPoint - BIセンターセミナー2013.pptx[読み取り専用]](/thumbs/94/120617602.jpg "Microsoft PowerPoint - BIセンターセミナー2013.pptx[読み取り専用]") 遺伝子配列解析の基礎 genome=gene+ome DNA 配列からタンパク質へ cgtgctttccacgacggtgacacgcttccctggattggccagactgccttccgggtcactgccatggaggagccgcagtcagatcctagcgtcgagccccctctga gtcaggaaacattttcagacctatggaaactacttcctgaaaacaacgttctgtcccccttgccgtcccaagcaatggatgatttgatgctgtccccggacgatattga
遺伝子配列解析の基礎 genome=gene+ome DNA 配列からタンパク質へ cgtgctttccacgacggtgacacgcttccctggattggccagactgccttccgggtcactgccatggaggagccgcagtcagatcctagcgtcgagccccctctga gtcaggaaacattttcagacctatggaaactacttcctgaaaacaacgttctgtcccccttgccgtcccaagcaatggatgatttgatgctgtccccggacgatattga
底膜は基板上にラミニン由来の接着ペプチドを結合させることにより, 基底膜の機能 を模倣した膜である. 応用例として皮膚移植が挙げられ, 移植医療などに利用される ことが期待されている. 2. 目的これまで, ラミニンから多くの細胞接着ペプチドが同定されているが, 上述した EF1 ペプチドは α2β
 氏名 ( 本籍 ) 山田寛尚 ( 東京都 ) 学位の種類博士 ( 生命科学 ) 学位記番号学位授与の日付学位授与の要件学位論文題目論文審査委員 博第 101 号平成 29 年 3 月 15 日学位規則第 4 条第 1 項該当分子動力学法を用いたラミニン由来ペプチドの構造解析 ( 主査 ) 高須昌子教授 小島正樹教授 野水基義教授 森本高子准教授 論文内容の要旨 1. 背景生体内を構成するオルガネラ,
氏名 ( 本籍 ) 山田寛尚 ( 東京都 ) 学位の種類博士 ( 生命科学 ) 学位記番号学位授与の日付学位授与の要件学位論文題目論文審査委員 博第 101 号平成 29 年 3 月 15 日学位規則第 4 条第 1 項該当分子動力学法を用いたラミニン由来ペプチドの構造解析 ( 主査 ) 高須昌子教授 小島正樹教授 野水基義教授 森本高子准教授 論文内容の要旨 1. 背景生体内を構成するオルガネラ,
1. 背景血小板上の受容体 CLEC-2 と ある種のがん細胞の表面に発現するタンパク質 ポドプラニン やマムシ毒 ロドサイチン が結合すると 血小板が活性化され 血液が凝固します ( 図 1) ポドプラニンは O- 結合型糖鎖が結合した糖タンパク質であり CLEC-2 受容体との結合にはその糖鎖が
 ポドプラニンは O- 結合型糖鎖が結合した糖タンパク質であり CLEC-2 受容体との結合にはその糖鎖が") 参考資料配布 2014 年 11 月 10 日 独立行政法人理化学研究所 国立大学法人東北大学 血小板上の受容体 CLEC-2 は糖鎖とペプチド鎖の両方を認識 - マムシ毒は糖鎖に依存せず受容体と結合 - 本研究成果のポイント レクチンは糖鎖とのみ結合する というこれまでの考え方を覆す CLEC-2 受容体は同じ領域でマムシ毒とがんに関わる糖タンパク質に結合 糖鎖を模倣したペプチド性薬剤の設計への応用に期待
参考資料配布 2014 年 11 月 10 日 独立行政法人理化学研究所 国立大学法人東北大学 血小板上の受容体 CLEC-2 は糖鎖とペプチド鎖の両方を認識 - マムシ毒は糖鎖に依存せず受容体と結合 - 本研究成果のポイント レクチンは糖鎖とのみ結合する というこれまでの考え方を覆す CLEC-2 受容体は同じ領域でマムシ毒とがんに関わる糖タンパク質に結合 糖鎖を模倣したペプチド性薬剤の設計への応用に期待
Microsoft PowerPoint - DNA1.ppt [互換モード]
![Microsoft PowerPoint - DNA1.ppt [互換モード]](/thumbs/94/118241118.jpg "Microsoft PowerPoint - DNA1.ppt [互換モード]") 生物物理化学 タンパク質をコードする遺伝子 (135~) 本 PPT 資料の作成には福岡大学機能生物研究室のホームページを参考にした http://133.100.212.50/~bc1/biochem/index2.htm 1 DA( デオキシリボ核酸 ) の化学的特徴 シャルガフ則とDAのX 線回折像をもとに,DAの構造が予測された (Watson & Crick 1953 年 ) 2 Watson
生物物理化学 タンパク質をコードする遺伝子 (135~) 本 PPT 資料の作成には福岡大学機能生物研究室のホームページを参考にした http://133.100.212.50/~bc1/biochem/index2.htm 1 DA( デオキシリボ核酸 ) の化学的特徴 シャルガフ則とDAのX 線回折像をもとに,DAの構造が予測された (Watson & Crick 1953 年 ) 2 Watson
人工知能補足_池村
 私くしにとって 生涯の指針となっている木村先生の教え 1. 想定外の発見の重要性 à unsupervised data mining for big data 2. 技術への信頼と技術開発の重要性 2D gel à BLSOM trna の二次元分離 : Methods in Enzymology 長さに依存する分離 想定外の 米国での Post Doc の時代 高分離能 長さに依存しない分離 29
私くしにとって 生涯の指針となっている木村先生の教え 1. 想定外の発見の重要性 à unsupervised data mining for big data 2. 技術への信頼と技術開発の重要性 2D gel à BLSOM trna の二次元分離 : Methods in Enzymology 長さに依存する分離 想定外の 米国での Post Doc の時代 高分離能 長さに依存しない分離 29
Microsoft PowerPoint マクロ生物学9
 マクロ生物学 9 生物は様々な化学反応で動いている 大阪大学工学研究科応用生物工学専攻細胞動態学領域 : 福井希一 1 生物の物質的基盤 Deleted based on copyright concern. カープ分子細胞生物学 より 2 8. 生物は様々な化学反応で動い ている 1. 生命の化学的基礎 2. 生命の物理法則 3 1. 生命の化学的基礎 1. 結合 2. 糖 脂質 3. 核酸 4.
マクロ生物学 9 生物は様々な化学反応で動いている 大阪大学工学研究科応用生物工学専攻細胞動態学領域 : 福井希一 1 生物の物質的基盤 Deleted based on copyright concern. カープ分子細胞生物学 より 2 8. 生物は様々な化学反応で動い ている 1. 生命の化学的基礎 2. 生命の物理法則 3 1. 生命の化学的基礎 1. 結合 2. 糖 脂質 3. 核酸 4.
の活性化が背景となるヒト悪性腫瘍の治療薬開発につながる 図4 研究である 研究内容 私たちは図3に示すようなyeast two hybrid 法を用いて AKT分子に結合する細胞内分子のスクリーニングを行った この結果 これまで機能の分からなかったプロトオンコジン TCL1がAKTと結合し多量体を形
 AKT活性を抑制するペプチ ド阻害剤の開発 野口 昌幸 北海道大学遺伝子病制御研究所 教授 広村 信 北海道大学遺伝子病制御研究所 ポスドク 岡田 太 北海道大学遺伝子病制御研究所 助手 柳舘 拓也 株式会社ラボ 研究員 ナーゼAKTに結合するタンパク分子を検索し これまで機能の 分からなかったプロトオンコジンTCL1がAKTと結合し AKT の活性化を促す AKT活性補助因子 であることを見い出し
AKT活性を抑制するペプチ ド阻害剤の開発 野口 昌幸 北海道大学遺伝子病制御研究所 教授 広村 信 北海道大学遺伝子病制御研究所 ポスドク 岡田 太 北海道大学遺伝子病制御研究所 助手 柳舘 拓也 株式会社ラボ 研究員 ナーゼAKTに結合するタンパク分子を検索し これまで機能の 分からなかったプロトオンコジンTCL1がAKTと結合し AKT の活性化を促す AKT活性補助因子 であることを見い出し
1 研究開発のねらい 糖鎖は 細胞表面のタンパク質や脂質に結合し 血液型の決定 細胞接着 抗原抗体反応 ウイルス感染などの生体反応で重要な役割を果たす生体分子である 糖鎖による多様な生物学的機能のうち 糖鎖結合タンパク質による糖鎖の特異的認識があり 糖鎖 - タンパク質間の相互作用の解析に糖鎖アレイ
 ライフサイエンスデータベース統合推進事業統合データ解析トライアル研究開発課題 タンパク質 - 糖鎖間の糖鎖結合部位の解明のためのツール改良及び解析 研究開発終了報告書 研究開発期間 : 平成 25 年 9 月 ~ 平成 26 年 1 月 研究代表者 : 細田正恵 ( 創価大学大学院工学研究科生命情報工学専攻 大学院生 ) - 1-2014 細田正恵 ( 創価大学大学院 )licensed under
ライフサイエンスデータベース統合推進事業統合データ解析トライアル研究開発課題 タンパク質 - 糖鎖間の糖鎖結合部位の解明のためのツール改良及び解析 研究開発終了報告書 研究開発期間 : 平成 25 年 9 月 ~ 平成 26 年 1 月 研究代表者 : 細田正恵 ( 創価大学大学院工学研究科生命情報工学専攻 大学院生 ) - 1-2014 細田正恵 ( 創価大学大学院 )licensed under
バイナリーパターン法によるデノボタンパク質WA20の
 配列パターンデザインによる新規人工設計タンパク質の半合理的創製と構造解析およびナノ構造構築への応用信州大学繊維学部応用生物科学系新井亮一 1. はじめに新規人工設計タンパク質 ( デノボタンパク質 de novo protein) とは 天然タンパク質のアミノ酸配列をもとにせず 新規にアミノ酸配列を設計した人工タンパク質である タンパク質を自由自在にデザインし 望みの構造や機能を創り出すことができるようになれば
配列パターンデザインによる新規人工設計タンパク質の半合理的創製と構造解析およびナノ構造構築への応用信州大学繊維学部応用生物科学系新井亮一 1. はじめに新規人工設計タンパク質 ( デノボタンパク質 de novo protein) とは 天然タンパク質のアミノ酸配列をもとにせず 新規にアミノ酸配列を設計した人工タンパク質である タンパク質を自由自在にデザインし 望みの構造や機能を創り出すことができるようになれば
生命情報学
 生命情報学 5 隠れマルコフモデル 阿久津達也 京都大学化学研究所 バイオインフォマティクスセンター 内容 配列モチーフ 最尤推定 ベイズ推定 M 推定 隠れマルコフモデル HMM Verアルゴリズム EMアルゴリズム Baum-Welchアルゴリズム 前向きアルゴリズム 後向きアルゴリズム プロファイル HMM 配列モチーフ モチーフ発見 配列モチーフ : 同じ機能を持つ遺伝子配列などに見られる共通の文字列パターン
生命情報学 5 隠れマルコフモデル 阿久津達也 京都大学化学研究所 バイオインフォマティクスセンター 内容 配列モチーフ 最尤推定 ベイズ推定 M 推定 隠れマルコフモデル HMM Verアルゴリズム EMアルゴリズム Baum-Welchアルゴリズム 前向きアルゴリズム 後向きアルゴリズム プロファイル HMM 配列モチーフ モチーフ発見 配列モチーフ : 同じ機能を持つ遺伝子配列などに見られる共通の文字列パターン
CourseDescriptions_
 MOE 2018.01 トレーニングコース説明 MOE の基本操作... 1 タンパク質の基本操作... 1 抗体設計... 2 タンパク質設計... 2 LBDD と SAR 解析... 3 SBDD とリガンド設計... 3 ファーマコフォア... 3 FBDD... 4 ケモインフォマティクスと QSAR... 4 SVL 基礎... 4 アドバンスト SBDD... 5 X 線結晶構造解析者向けツール...
MOE 2018.01 トレーニングコース説明 MOE の基本操作... 1 タンパク質の基本操作... 1 抗体設計... 2 タンパク質設計... 2 LBDD と SAR 解析... 3 SBDD とリガンド設計... 3 ファーマコフォア... 3 FBDD... 4 ケモインフォマティクスと QSAR... 4 SVL 基礎... 4 アドバンスト SBDD... 5 X 線結晶構造解析者向けツール...
1_alignment.ppt
 " " " " n " n n " n " n n n " n n n n " n LGPSSKQTGKGW-SRIWDN! + +! LN-ITKSAGKGAIMRLGDA! " n -------TGKG--------!! -------AGKG--------! " n w w w " n w w " " " 11 12 " n w w w " n w w A! M! O! A!
" " " " n " n n " n " n n n " n n n n " n LGPSSKQTGKGW-SRIWDN! + +! LN-ITKSAGKGAIMRLGDA! " n -------TGKG--------!! -------AGKG--------! " n w w w " n w w " " " 11 12 " n w w w " n w w A! M! O! A!
Microsoft PowerPoint - 3rd-jikken-vscreen [互換モード]
![Microsoft PowerPoint - 3rd-jikken-vscreen [互換モード]](/thumbs/95/123159298.jpg "Microsoft PowerPoint - 3rd-jikken-vscreen [互換モード]") 生命情報実験第一 ( 情報系 ) バイオインフォマティクスの道具箱 タンパク質化合物相互作用解析: バーチャルスクリーニング 慶應義塾大学生命情報学科榊原康文, 佐藤健吾 リード化合物探索とインフォマティクス High Throughput Screening 実験的検証 リード化合物 = 薬剤候補 薬剤標的タンパク質 初期候補 実験的検証 + インフォマティクス 1. 大量化合物の探索 2. 成功率向上
生命情報実験第一 ( 情報系 ) バイオインフォマティクスの道具箱 タンパク質化合物相互作用解析: バーチャルスクリーニング 慶應義塾大学生命情報学科榊原康文, 佐藤健吾 リード化合物探索とインフォマティクス High Throughput Screening 実験的検証 リード化合物 = 薬剤候補 薬剤標的タンパク質 初期候補 実験的検証 + インフォマティクス 1. 大量化合物の探索 2. 成功率向上

受精に関わる精子融合因子 IZUMO1 と卵子受容体 JUNO の認識機構を解明 1. 発表者 : 大戸梅治 ( 東京大学大学院薬学系研究科准教授 ) 石田英子 ( 東京大学大学院薬学系研究科特任研究員 ) 清水敏之 ( 東京大学大学院薬学系研究科教授 ) 井上直和 ( 福島県立医科大学医学部附属生
 石田英子 ( 東京大学大学院薬学系研究科特任研究員 ) 清水敏之 ( 東京大学大学院薬学系研究科教授 ) 井上直和 ( 福島県立医科大学医学部附属生") 受精に関わる精子融合因子 IZUMO1 と卵子受容体 JUNO の認識機構を解明 1. 発表者 : 大戸梅治 ( 東京大学大学院薬学系研究科准教授 ) 石田英子 ( 東京大学大学院薬学系研究科特任研究員 ) 清水敏之 ( 東京大学大学院薬学系研究科教授 ) 井上直和 ( 福島県立医科大学医学部附属生体情報伝達研究所准教授 ) 内山進 ( 大阪大学大学院工学研究科准教授 ) 2. 発表のポイント :
受精に関わる精子融合因子 IZUMO1 と卵子受容体 JUNO の認識機構を解明 1. 発表者 : 大戸梅治 ( 東京大学大学院薬学系研究科准教授 ) 石田英子 ( 東京大学大学院薬学系研究科特任研究員 ) 清水敏之 ( 東京大学大学院薬学系研究科教授 ) 井上直和 ( 福島県立医科大学医学部附属生体情報伝達研究所准教授 ) 内山進 ( 大阪大学大学院工学研究科准教授 ) 2. 発表のポイント :
Microsoft Word - レポート模範例.docx
 生物学レポート ヒトとウシのヘモグロビン α 鎖と酵素タンパク質 trypsin について 一次構造と高次構造を調べ比較する 畜産学科 1 年学籍番号 00000000 氏名藍愛子 提出日 :2012 年 6 月 27 日 1 はじめに レポートの課題説明に記述されていた このレポートの課題は以下のように要約される 大学でそれぞれの科目を学習する大きな目的と意義は ある学問体系の中で 何がわかっていて
生物学レポート ヒトとウシのヘモグロビン α 鎖と酵素タンパク質 trypsin について 一次構造と高次構造を調べ比較する 畜産学科 1 年学籍番号 00000000 氏名藍愛子 提出日 :2012 年 6 月 27 日 1 はじめに レポートの課題説明に記述されていた このレポートの課題は以下のように要約される 大学でそれぞれの科目を学習する大きな目的と意義は ある学問体系の中で 何がわかっていて
 Structure クイックスタート このミニコースでは NCBI から提供されている Cn3D を用いてどのようにタンパク質の立体構造を可視化や注釈付けを行うか さらにはタンパク質内で保存されているドメインの確認の方法 同様のドメインを持っている他のタンパク質の探索方法 3D モデリングのテンプレートとなるタンパク質の見つけ方 BLAST では認識できないような非常に離れたホモログの検出方法について学びます
Structure クイックスタート このミニコースでは NCBI から提供されている Cn3D を用いてどのようにタンパク質の立体構造を可視化や注釈付けを行うか さらにはタンパク質内で保存されているドメインの確認の方法 同様のドメインを持っている他のタンパク質の探索方法 3D モデリングのテンプレートとなるタンパク質の見つけ方 BLAST では認識できないような非常に離れたホモログの検出方法について学びます
バイオインフォマティクスⅠ
 バイオインフォマティクス ( 第 5 回 ) 慶應義塾大学生命情報学科 榊原康文 多重アライメントの解 0 2 3 4 5 6 7 j Q T S Y T R Y Q T - Y T R K 0 0-9 -20-44 -52-63 -72-90 Q -6 2 0-6 -4-25 -34-52 2 S -32 5 30 4 6-5 -4-32 3 Y -48-4 2 38 27 8 0 4 P -64-27
バイオインフォマティクス ( 第 5 回 ) 慶應義塾大学生命情報学科 榊原康文 多重アライメントの解 0 2 3 4 5 6 7 j Q T S Y T R Y Q T - Y T R K 0 0-9 -20-44 -52-63 -72-90 Q -6 2 0-6 -4-25 -34-52 2 S -32 5 30 4 6-5 -4-32 3 Y -48-4 2 38 27 8 0 4 P -64-27
<4D F736F F D20838C837C815B836789DB91E890E096BE2E646F6378>
 レポートについて 1. 課題 以下に記した手順に従って ヒトのヘモグロビンα 鎖タンパク質と酵素タンパク質 trypsin について その一次構造をタンパク質データベースにアクセスして調べ さらにその二次構造と三次構造を ProteinDataBank へアクセスして確認する 以上の経過と結果を いつ どこで調べたかを含めてその過程を記述し さらに検索結果である両タンパク質の一次構造 分子の形 (
レポートについて 1. 課題 以下に記した手順に従って ヒトのヘモグロビンα 鎖タンパク質と酵素タンパク質 trypsin について その一次構造をタンパク質データベースにアクセスして調べ さらにその二次構造と三次構造を ProteinDataBank へアクセスして確認する 以上の経過と結果を いつ どこで調べたかを含めてその過程を記述し さらに検索結果である両タンパク質の一次構造 分子の形 (
生物時計の安定性の秘密を解明
 平成 25 年 12 月 13 日 生物時計の安定性の秘密を解明 概要 名古屋大学理学研究科の北山陽子助教 近藤孝男特任教授らの研究グループは 光合 成をおこなうシアノバクテリアの生物時計機構を解析し 時計タンパク質 KaiC が 安定な 24 時 間周期のリズムを形成する分子機構を明らかにしました 生物は, 生物時計 ( 概日時計 ) を利用して様々な生理現象を 時間的に コントロールし 効 率的に生活しています
平成 25 年 12 月 13 日 生物時計の安定性の秘密を解明 概要 名古屋大学理学研究科の北山陽子助教 近藤孝男特任教授らの研究グループは 光合 成をおこなうシアノバクテリアの生物時計機構を解析し 時計タンパク質 KaiC が 安定な 24 時 間周期のリズムを形成する分子機構を明らかにしました 生物は, 生物時計 ( 概日時計 ) を利用して様々な生理現象を 時間的に コントロールし 効 率的に生活しています
バイオインフォマティクスⅠ
 バイオインフォマティクス ( 第 3 回 ) 慶應義塾大学生命情報学科 榊原康文 アセンブリの演習問題 ( 解 ) CGTCCGT CATCG 5 3 4 ATCCAT TCCGTAT 5 3 3 4 GTATC CGTCCGT-------- --TCCGTAT------ -----GTATC----- -------ATCCAT-- ----------CATCG ===============
バイオインフォマティクス ( 第 3 回 ) 慶應義塾大学生命情報学科 榊原康文 アセンブリの演習問題 ( 解 ) CGTCCGT CATCG 5 3 4 ATCCAT TCCGTAT 5 3 3 4 GTATC CGTCCGT-------- --TCCGTAT------ -----GTATC----- -------ATCCAT-- ----------CATCG ===============
 1 911 9001030 9:00 A B C D E F G H I J K L M 1A0900 1B0900 1C0900 1D0900 1E0900 1F0900 1G0900 1H0900 1I0900 1J0900 1K0900 1L0900 1M0900 9:15 1A0915 1B0915 1C0915 1D0915 1E0915 1F0915 1G0915 1H0915 1I0915
1 911 9001030 9:00 A B C D E F G H I J K L M 1A0900 1B0900 1C0900 1D0900 1E0900 1F0900 1G0900 1H0900 1I0900 1J0900 1K0900 1L0900 1M0900 9:15 1A0915 1B0915 1C0915 1D0915 1E0915 1F0915 1G0915 1H0915 1I0915
問 1 正解 2 大腸菌 bp ショウジョウバエ bp ヒト bp パンコムギ bp 問 2 正解 4 遺伝子地図には組換え率から得られる連鎖地図と物理的距離に基づく物理地図とがあり その作成においては SNP RFLP マイ
 平成 25 年度日本バイオインフォマティクス学会 (JSBi) バイオインフォマティクス技術者認定試験解説集 - 1 - 問 1 正解 2 大腸菌 4.6 10 6 bp ショウジョウバエ 1.8 10 8 bp ヒト 3.0 10 9 bp パンコムギ 1.7 10 10 bp 問 2 正解 4 遺伝子地図には組換え率から得られる連鎖地図と物理的距離に基づく物理地図とがあり その作成においては SNP
平成 25 年度日本バイオインフォマティクス学会 (JSBi) バイオインフォマティクス技術者認定試験解説集 - 1 - 問 1 正解 2 大腸菌 4.6 10 6 bp ショウジョウバエ 1.8 10 8 bp ヒト 3.0 10 9 bp パンコムギ 1.7 10 10 bp 問 2 正解 4 遺伝子地図には組換え率から得られる連鎖地図と物理的距離に基づく物理地図とがあり その作成においては SNP
報道発表資料 2007 年 8 月 1 日 独立行政法人理化学研究所 マイクロ RNA によるタンパク質合成阻害の仕組みを解明 - mrna の翻訳が抑制される過程を試験管内で再現することに成功 - ポイント マイクロ RNA が翻訳の開始段階を阻害 標的 mrna の尻尾 ポリ A テール を短縮
 60 秒でわかるプレスリリース 2007 年 8 月 1 日 独立行政法人理化学研究所 マイクロ RNA によるタンパク質合成阻害の仕組みを解明 - mrna の翻訳が抑制される過程を試験管内で再現することに成功 - 生命は 遺伝子の設計図をもとにつくられるタンパク質によって 営まれています タンパク質合成は まず DNA 情報がいったん mrna に転写され 次に mrna がタンパク質の合成工場である
60 秒でわかるプレスリリース 2007 年 8 月 1 日 独立行政法人理化学研究所 マイクロ RNA によるタンパク質合成阻害の仕組みを解明 - mrna の翻訳が抑制される過程を試験管内で再現することに成功 - 生命は 遺伝子の設計図をもとにつくられるタンパク質によって 営まれています タンパク質合成は まず DNA 情報がいったん mrna に転写され 次に mrna がタンパク質の合成工場である
平成20年度 神戸大学 大学院理学研究科 化学専攻 入学試験問題
 化学 Ⅰ- 表紙 平成 31 年度神戸大学大学院理学研究科化学専攻入学試験 化学 Ⅰ 試験時間 10:30-11:30(60 分 ) 表紙を除いて 7 ページあります 問題 [Ⅰ]~ 問題 [Ⅵ] の中から 4 題を選択して 解答しなさい 各ページ下端にある 選択する 選択しない のうち 該当する方を丸で囲みなさい 各ページに ( 用紙上端 ) と ( 用紙下端 ) を記入しなさい を誤って記入すると採点の対象とならないことがあります
化学 Ⅰ- 表紙 平成 31 年度神戸大学大学院理学研究科化学専攻入学試験 化学 Ⅰ 試験時間 10:30-11:30(60 分 ) 表紙を除いて 7 ページあります 問題 [Ⅰ]~ 問題 [Ⅵ] の中から 4 題を選択して 解答しなさい 各ページ下端にある 選択する 選択しない のうち 該当する方を丸で囲みなさい 各ページに ( 用紙上端 ) と ( 用紙下端 ) を記入しなさい を誤って記入すると採点の対象とならないことがあります
5_motif 公開版.ppt
 配列モチーフ 機能ドメイン 機能部位 機能的 構造的に重要な部位 は進化の過程で保存 される傾向がある 進化的に保存された ドメイン 配列モチーフ 機能ドメイン中の特徴的な 保存配列パターン マルチプルアライメント から抽出 配列モチーフの表現方法 パターン プロファイル 2 n n n n n n n n ENCODE n PROSITE パターンの例 n C-x(2,4)-C-x(3)-[LIVMFYWC]-x(8)-H-x(3,5)-H.
配列モチーフ 機能ドメイン 機能部位 機能的 構造的に重要な部位 は進化の過程で保存 される傾向がある 進化的に保存された ドメイン 配列モチーフ 機能ドメイン中の特徴的な 保存配列パターン マルチプルアライメント から抽出 配列モチーフの表現方法 パターン プロファイル 2 n n n n n n n n ENCODE n PROSITE パターンの例 n C-x(2,4)-C-x(3)-[LIVMFYWC]-x(8)-H-x(3,5)-H.
細胞骨格を形成するタンパク質
 タンパク質に関する練習問題総まとめ 第 1 回タンパク質の一次構造と高次構造 1. ペプチド結合部分は平面構造を取る 2. 分子シャペロンは タンパク質の ( ) をする働きをもつ 3. 動物種が違っても 同じ作用を持つタンパク質は 同一のアミノ酸配列からなる 4. ある 2 つのタンパク質間の相同性 (%) は それらのホモロジー (%) より高い 5. 加熱処理により タンパク質を安定化している
タンパク質に関する練習問題総まとめ 第 1 回タンパク質の一次構造と高次構造 1. ペプチド結合部分は平面構造を取る 2. 分子シャペロンは タンパク質の ( ) をする働きをもつ 3. 動物種が違っても 同じ作用を持つタンパク質は 同一のアミノ酸配列からなる 4. ある 2 つのタンパク質間の相同性 (%) は それらのホモロジー (%) より高い 5. 加熱処理により タンパク質を安定化している
<4D F736F F D B82C982C282A282C482512E646F63>
 サンプル条件および固定化分子の選択 Biacoreの実験ではセンサーチップに固定化する分子をリガンド それに対して結合を測定する分子をアナライトと呼びます いずれの分子をリガンドとし アナライトとするかは 実験系を構築する上で重要です 以下にサンプルに適したリガンド アナライトの設計方法やサンプルの必要条件などをご紹介します アナライト リガンド センサーチップ (1) タンパク質リガンドとしてもアナライトとしても用いることができます
サンプル条件および固定化分子の選択 Biacoreの実験ではセンサーチップに固定化する分子をリガンド それに対して結合を測定する分子をアナライトと呼びます いずれの分子をリガンドとし アナライトとするかは 実験系を構築する上で重要です 以下にサンプルに適したリガンド アナライトの設計方法やサンプルの必要条件などをご紹介します アナライト リガンド センサーチップ (1) タンパク質リガンドとしてもアナライトとしても用いることができます
FMO法のリガンド-タンパク質相互作用解析への応用
 2010 年 10 月 8 日学術総合センター 地球シミュレータ産業利用シンポジウム 2010 フラグメント分子軌道 (FMO) 法の創薬に おける分子シミュレーションへの応用 小澤基裕 * 1 小沢知永 * 1 半田千彰 * 1 辻英一 * 1 岡崎浩輔 * 1 新宮哲 * 2 数納広哉 * 2 上原均 * 2 *1 キッセイ薬品工業株式会社創薬研究部創薬基盤研究所 *2 独立行政法人海洋研究開発機構
2010 年 10 月 8 日学術総合センター 地球シミュレータ産業利用シンポジウム 2010 フラグメント分子軌道 (FMO) 法の創薬に おける分子シミュレーションへの応用 小澤基裕 * 1 小沢知永 * 1 半田千彰 * 1 辻英一 * 1 岡崎浩輔 * 1 新宮哲 * 2 数納広哉 * 2 上原均 * 2 *1 キッセイ薬品工業株式会社創薬研究部創薬基盤研究所 *2 独立行政法人海洋研究開発機構
A Constructive Approach to Gene Expression Dynamics
 配列アラインメント (I): 大域アラインメント http://www.lab.tohou.ac.jp/sci/is/nacher/eaching/bioinformatics/ week.pdf 08/4/0 08/4/0 基本的な考え方 バイオインフォマティクスにはさまざまなアルゴリズムがありますが その多くにおいて基本的な考え方は 配列が類似していれば 機能も類似している というものである 例えば
配列アラインメント (I): 大域アラインメント http://www.lab.tohou.ac.jp/sci/is/nacher/eaching/bioinformatics/ week.pdf 08/4/0 08/4/0 基本的な考え方 バイオインフォマティクスにはさまざまなアルゴリズムがありますが その多くにおいて基本的な考え方は 配列が類似していれば 機能も類似している というものである 例えば
Microsoft PowerPoint - BI_okuno_
 バイオインフォマティクス ( 配列検索 ) & ケモインフォマティクス ( 構造検索 ) 統合薬学教育開発分野 奥野恭史 創薬におけるインフォマティクス ゲノム情報 ゲノム基盤ターゲット研究探索 ターゲット バリデーション 創薬リード探索 創薬リード最適化 前臨床研究臨床研究 創薬 ゲノム情報 (~2 万 2 千遺伝子 ) 化合物ライブラリー (10^60 化合物 ) バイオインフォマティクス ケモインフォマティクス
バイオインフォマティクス ( 配列検索 ) & ケモインフォマティクス ( 構造検索 ) 統合薬学教育開発分野 奥野恭史 創薬におけるインフォマティクス ゲノム情報 ゲノム基盤ターゲット研究探索 ターゲット バリデーション 創薬リード探索 創薬リード最適化 前臨床研究臨床研究 創薬 ゲノム情報 (~2 万 2 千遺伝子 ) 化合物ライブラリー (10^60 化合物 ) バイオインフォマティクス ケモインフォマティクス
Slide 1
 転写 1. タンパク合成における RNA の役割酵素誘導 2. RNA ポリメラーゼ鎖型への結合転写開始鎖延長転写終結真核生物の RNA ポリメラーゼ 3. 原核生物における転写制御プロモーターカタボライト ( 異化代謝産物 ) 抑制オペロン 4. 転写後修飾プロセシング RNA ポリメラーゼ ( 鎖型への結合 ) プロモーターに特異的に結合 大腸菌の代表的なプロモーターのセンス鎖の配列 RNA ポリメラーゼ
転写 1. タンパク合成における RNA の役割酵素誘導 2. RNA ポリメラーゼ鎖型への結合転写開始鎖延長転写終結真核生物の RNA ポリメラーゼ 3. 原核生物における転写制御プロモーターカタボライト ( 異化代謝産物 ) 抑制オペロン 4. 転写後修飾プロセシング RNA ポリメラーゼ ( 鎖型への結合 ) プロモーターに特異的に結合 大腸菌の代表的なプロモーターのセンス鎖の配列 RNA ポリメラーゼ
「組換えDNA技術応用食品及び添加物の安全性審査の手続」の一部改正について
 食安基発 0627 第 3 号 平成 26 年 6 月 27 日 各検疫所長殿 医薬食品局食品安全部基準審査課長 ( 公印省略 ) 最終的に宿主に導入されたDNAが 当該宿主と分類学上同一の種に属する微生物のDNAのみである場合又は組換え体が自然界に存在する微生物と同等の遺伝子構成である場合のいずれかに該当することが明らかであると判断する基準に係る留意事項について 食品 添加物等の規格基準 ( 昭和
食安基発 0627 第 3 号 平成 26 年 6 月 27 日 各検疫所長殿 医薬食品局食品安全部基準審査課長 ( 公印省略 ) 最終的に宿主に導入されたDNAが 当該宿主と分類学上同一の種に属する微生物のDNAのみである場合又は組換え体が自然界に存在する微生物と同等の遺伝子構成である場合のいずれかに該当することが明らかであると判断する基準に係る留意事項について 食品 添加物等の規格基準 ( 昭和
バイオインフォマティクスⅠ
 ポストゲノム生命科学方法論 榊原担当の第 回 慶應義塾大学生命情報学科 榊原康文 遺伝子発現プロファイルの解析手法 クラスタリング Unsupervsed 階層クラスタリング,k-means 法, 自己組織化マップ SOM 識別 Supervsed 線形識別関数,k- 近傍法, サポートベクターマシン ブール関数, ニューラルネットワーク 遺伝子発現データの識別 マイクロアレイの遺伝子発現データを
ポストゲノム生命科学方法論 榊原担当の第 回 慶應義塾大学生命情報学科 榊原康文 遺伝子発現プロファイルの解析手法 クラスタリング Unsupervsed 階層クラスタリング,k-means 法, 自己組織化マップ SOM 識別 Supervsed 線形識別関数,k- 近傍法, サポートベクターマシン ブール関数, ニューラルネットワーク 遺伝子発現データの識別 マイクロアレイの遺伝子発現データを
Powered by TCPDF ( Title ハイリスクHPV 型のタンパクを標的とした新たな分子標的治療に関する基礎的検討 Sub Title Study of a new therapy targeting proteins of high-risk HPV Au
 Powered by TCPDF (www.tcpdf.org) Title ハイリスクHPV 型のタンパクを標的とした新たな分子標的治療に関する基礎的検討 Sub Title Study of a new therapy targeting proteins of high-risk HPV Author 大野, 暁子 (Ono, Akiko) Publisher Publication year
Powered by TCPDF (www.tcpdf.org) Title ハイリスクHPV 型のタンパクを標的とした新たな分子標的治療に関する基礎的検討 Sub Title Study of a new therapy targeting proteins of high-risk HPV Author 大野, 暁子 (Ono, Akiko) Publisher Publication year
分子系統解析における様々な問題について 田辺晶史
 分子系統解析における様々な問題について 田辺晶史 そもそもどこの配列を使うべき? そもそもどこの配列を使うべき? 置換が早すぎず遅すぎない (= 多すぎず少なすぎない ) そもそもどこの配列を使うべき? 置換が早すぎず遅すぎない (= 多すぎず少なすぎない ) 連続長は長い方が良い そもそもどこの配列を使うべき? 置換が早すぎず遅すぎない (= 多すぎず少なすぎない ) 連続長は長い方が良い 遺伝子重複が起きていない
分子系統解析における様々な問題について 田辺晶史 そもそもどこの配列を使うべき? そもそもどこの配列を使うべき? 置換が早すぎず遅すぎない (= 多すぎず少なすぎない ) そもそもどこの配列を使うべき? 置換が早すぎず遅すぎない (= 多すぎず少なすぎない ) 連続長は長い方が良い そもそもどこの配列を使うべき? 置換が早すぎず遅すぎない (= 多すぎず少なすぎない ) 連続長は長い方が良い 遺伝子重複が起きていない
研究成果報告書
 様式 C-19 科学研究費補助金研究成果報告書 研究種目 : 基盤研究 (C) 研究期間 :2007 2008 課題番号 :19570157 研究課題名 ( 和文 ) 生体高分子間相互作用構造推定のため情報抽出法の開発 平成 21 年 5 月 31 日現在 研究課題名 ( 英文 )Development of a method to obtain information for prediction
様式 C-19 科学研究費補助金研究成果報告書 研究種目 : 基盤研究 (C) 研究期間 :2007 2008 課題番号 :19570157 研究課題名 ( 和文 ) 生体高分子間相互作用構造推定のため情報抽出法の開発 平成 21 年 5 月 31 日現在 研究課題名 ( 英文 )Development of a method to obtain information for prediction
れており 世界的にも重要課題とされています それらの中で 非常に高い完全長 cdna のカバー率を誇るマウスエンサイクロペディア計画は極めて重要です ゲノム科学総合研究センター (GSC) 遺伝子構造 機能研究グループでは これまでマウス完全長 cdna100 万クローン以上の末端塩基配列データを
 遺伝子構造 機能研究グループでは これまでマウス完全長 cdna100 万クローン以上の末端塩基配列データを") 報道発表資料 2002 年 12 月 5 日 独立行政法人理化学研究所 遺伝子の機能解析を飛躍的に進める世界最大規模の遺伝子情報を公開 - 遺伝子として認知されていなかった部分が転写されていることを実証 - 理化学研究所 ( 小林俊一理事長 ) は マウスの完全長 cdna 160,770 クローンの塩基配列および機能アノテーション ( 機能注釈 ) 情報を公開します これは 現在までに人類が収得している遺伝子の約
報道発表資料 2002 年 12 月 5 日 独立行政法人理化学研究所 遺伝子の機能解析を飛躍的に進める世界最大規模の遺伝子情報を公開 - 遺伝子として認知されていなかった部分が転写されていることを実証 - 理化学研究所 ( 小林俊一理事長 ) は マウスの完全長 cdna 160,770 クローンの塩基配列および機能アノテーション ( 機能注釈 ) 情報を公開します これは 現在までに人類が収得している遺伝子の約
「組換えDNA技術応用食品及び添加物の安全性審査の手続」の一部改正について
 ( 別添 ) 最終的に宿主に導入された DNA が 当該宿主と分類学上同一の種に属する微生物の DNA のみである場合又は組換え体が自然界に存在する微生物と同等の遺伝子構成である場合のいずれかに該当することが明らかであると判断する基準に係る留意事項 最終的に宿主に導入されたDNAが 当該宿主と分類学上同一の種に属する微生物のDNAのみである場合又は組換え体が自然界に存在する微生物と同等の遺伝子構成である場合のいずれかに該当することが明らかであると判断する基準
( 別添 ) 最終的に宿主に導入された DNA が 当該宿主と分類学上同一の種に属する微生物の DNA のみである場合又は組換え体が自然界に存在する微生物と同等の遺伝子構成である場合のいずれかに該当することが明らかであると判断する基準に係る留意事項 最終的に宿主に導入されたDNAが 当該宿主と分類学上同一の種に属する微生物のDNAのみである場合又は組換え体が自然界に存在する微生物と同等の遺伝子構成である場合のいずれかに該当することが明らかであると判断する基準
ヒト脂肪組織由来幹細胞における外因性脂肪酸結合タンパク (FABP)4 FABP 5 の影響 糖尿病 肥満の病態解明と脂肪幹細胞再生治療への可能性 ポイント 脂肪幹細胞の脂肪分化誘導に伴い FABP4( 脂肪細胞型 ) FABP5( 表皮型 ) が発現亢進し 分泌されることを確認しました トランスク
4 FABP 5 の影響 糖尿病 肥満の病態解明と脂肪幹細胞再生治療への可能性 ポイント 脂肪幹細胞の脂肪分化誘導に伴い FABP4( 脂肪細胞型 ) FABP5( 表皮型 ) が発現亢進し 分泌されることを確認しました トランスク") 平成 28 年 12 月 19 日 ヒト脂肪組織由来幹細胞における外因性脂肪酸結合タンパク (FABP)4 FABP 5 の影響 糖尿病 肥満の病態解明と脂肪幹細胞再生治療への可能性 名古屋大学大学院医学系研究科 ( 研究科長 髙橋雅英 ) 泌尿器科学分野の山本徳則 ( やまもととくのり ) 准教授 後藤百万 ( ごとうももかず ) 教授と札幌医科大学内分泌内科の古橋眞人 ( ふるはしまさと ) 講師
平成 28 年 12 月 19 日 ヒト脂肪組織由来幹細胞における外因性脂肪酸結合タンパク (FABP)4 FABP 5 の影響 糖尿病 肥満の病態解明と脂肪幹細胞再生治療への可能性 名古屋大学大学院医学系研究科 ( 研究科長 髙橋雅英 ) 泌尿器科学分野の山本徳則 ( やまもととくのり ) 准教授 後藤百万 ( ごとうももかず ) 教授と札幌医科大学内分泌内科の古橋眞人 ( ふるはしまさと ) 講師
東京理科大学 Ⅰ 部化学研究部 2015 年度春輪講書 シクロデキストリンを用いた 包接化合物の生成 水曜班 Ikemura, M.(2C),Ebihara, K.(2C), Kataoka, T.(2K), Shibasaki,K.(2OK),Tsumeda,T.(2C),Naka,A.(2OK)
,Ebihara, K.(2C), Kataoka, T.(2K), Shibasaki,K.(2OK),Tsumeda,T.(2C),Naka,A.(2OK)") 東京理科大学 Ⅰ 部化学研究部 2015 年度春輪講書 シクロデキストリンを用いた 包接化合物の生成 水曜班 Ikemura, M.(2C),Ebihara, K.(2C), Kataoka, T.(2K), Shibasaki,K.(2OK),Tsumeda,T.(2C),Naka,A.(2OK),Murakoshi,R.(2OK), Okawa,T.(2C),Noguchi,A.(2K),Sekiguchi,K.(2K),Hashimoto,Y.(2C)
東京理科大学 Ⅰ 部化学研究部 2015 年度春輪講書 シクロデキストリンを用いた 包接化合物の生成 水曜班 Ikemura, M.(2C),Ebihara, K.(2C), Kataoka, T.(2K), Shibasaki,K.(2OK),Tsumeda,T.(2C),Naka,A.(2OK),Murakoshi,R.(2OK), Okawa,T.(2C),Noguchi,A.(2K),Sekiguchi,K.(2K),Hashimoto,Y.(2C)
図 B 細胞受容体を介した NF-κB 活性化モデル
 60 秒でわかるプレスリリース 2007 年 12 月 17 日 独立行政法人理化学研究所 免疫の要 NF-κB の活性化シグナルを増幅する機構を発見 - リン酸化酵素 IKK が正のフィーッドバックを担当 - 身体に病原菌などの異物 ( 抗原 ) が侵入すると 誰にでも備わっている免疫システムが働いて 異物を認識し 排除するために さまざまな反応を起こします その一つに 免疫細胞である B 細胞が
60 秒でわかるプレスリリース 2007 年 12 月 17 日 独立行政法人理化学研究所 免疫の要 NF-κB の活性化シグナルを増幅する機構を発見 - リン酸化酵素 IKK が正のフィーッドバックを担当 - 身体に病原菌などの異物 ( 抗原 ) が侵入すると 誰にでも備わっている免疫システムが働いて 異物を認識し 排除するために さまざまな反応を起こします その一つに 免疫細胞である B 細胞が
Microsoft PowerPoint - lecture a.pptx
 応用生命科学 情報生命学第 3 回配列解析入門 7 月 14 日 ( 木 ) 3 時限目加藤有己大阪大学大学院医学系研究科講義資料 http://www.med.osakau.ac.p/pub/rna/ykato/lecture/bonfo16/ 授業目的 情報科学と生命科学の融合領域である情報生命科学の基本的な手法を理解することを目的とする 日程 3 時限目 4 時限目 6 月 30 日 ( 木
応用生命科学 情報生命学第 3 回配列解析入門 7 月 14 日 ( 木 ) 3 時限目加藤有己大阪大学大学院医学系研究科講義資料 http://www.med.osakau.ac.p/pub/rna/ykato/lecture/bonfo16/ 授業目的 情報科学と生命科学の融合領域である情報生命科学の基本的な手法を理解することを目的とする 日程 3 時限目 4 時限目 6 月 30 日 ( 木
大学院博士課程共通科目ベーシックプログラム
 平成 30 年度医科学専攻共通科目 共通基礎科目実習 ( 旧コア実習 ) 概要 1 ). 大学院生が所属する教育研究分野における実習により単位認定可能な実習項目 ( コア実習項目 ) 1. 組換え DNA 技術実習 2. 生体物質の調製と解析実習 3. 薬理学実習 4. ウイルス学実習 5. 免疫学実習 6. 顕微鏡試料作成法実習 7. ゲノム医学実習 8. 共焦点レーザー顕微鏡実習 2 ). 実習を担当する教育研究分野においてのみ単位認定可能な実習項目
平成 30 年度医科学専攻共通科目 共通基礎科目実習 ( 旧コア実習 ) 概要 1 ). 大学院生が所属する教育研究分野における実習により単位認定可能な実習項目 ( コア実習項目 ) 1. 組換え DNA 技術実習 2. 生体物質の調製と解析実習 3. 薬理学実習 4. ウイルス学実習 5. 免疫学実習 6. 顕微鏡試料作成法実習 7. ゲノム医学実習 8. 共焦点レーザー顕微鏡実習 2 ). 実習を担当する教育研究分野においてのみ単位認定可能な実習項目
共起関係解析によるタンパク質の機能モジュール探索法の開発
 2013.11.29 統合データ解析トライアル 中間激励会 共起関係解析によるタンパク質の機能モジュール探索法の開発 九州工業大学 情報工 生命情報 藤井聡 2013 藤井聡 ( 九州工業大学 )licensed under CC 表示 2.1 日本 1 背景 疾病関連遺伝子 ドメイン モチーフ ドラッグターゲット T/G Gefitinib LUNG CANCER OMIM NHGRI GWAS
2013.11.29 統合データ解析トライアル 中間激励会 共起関係解析によるタンパク質の機能モジュール探索法の開発 九州工業大学 情報工 生命情報 藤井聡 2013 藤井聡 ( 九州工業大学 )licensed under CC 表示 2.1 日本 1 背景 疾病関連遺伝子 ドメイン モチーフ ドラッグターゲット T/G Gefitinib LUNG CANCER OMIM NHGRI GWAS
Microsoft PowerPoint - 3Dstructure08Jun3 [互換モード]
![Microsoft PowerPoint - 3Dstructure08Jun3 [互換モード]](/thumbs/102/156788389.jpg "Microsoft PowerPoint - 3Dstructure08Jun3 [互換モード]") 近畿大学 農学部 生命情報学 生体高分子の立体構造とその予測 Ras MTEYKLVVVGAGGVGKSAL TIQLIQHFVDEYDPTIED SYRKQVVIDGETLLDILD TAGQEEYSAMRDQYMRTGE GFLVFAITKSFEDIHQ YREQIKRVKDSDDVPMVLV GKDLAARTVESRQAQDL ARSYGIPYIETSAKTRQGV EDAFYTLVREIRQH
近畿大学 農学部 生命情報学 生体高分子の立体構造とその予測 Ras MTEYKLVVVGAGGVGKSAL TIQLIQHFVDEYDPTIED SYRKQVVIDGETLLDILD TAGQEEYSAMRDQYMRTGE GFLVFAITKSFEDIHQ YREQIKRVKDSDDVPMVLV GKDLAARTVESRQAQDL ARSYGIPYIETSAKTRQGV EDAFYTLVREIRQH
研究成果報告書
 様式 C-19 F-19 Z-19( 共通 ) 1. 研究開始当初の背景研究開始当初の平成 24 年の時点で HUGO (Human Genome Organisation) Gene Nomenculature Committee (http://www. genename.org) には ヒトの遺伝子記号が 32,000 登録されておりそのうちタンパク質をコードするものは約 19,000 であった
様式 C-19 F-19 Z-19( 共通 ) 1. 研究開始当初の背景研究開始当初の平成 24 年の時点で HUGO (Human Genome Organisation) Gene Nomenculature Committee (http://www. genename.org) には ヒトの遺伝子記号が 32,000 登録されておりそのうちタンパク質をコードするものは約 19,000 であった
木村の理論化学小ネタ 液体と液体の混合物 ( 二成分系 ) の気液平衡 はじめに 純物質 A( 液体 ) と純物質 B( 液体 ) が存在し, 分子 A の間に働く力 分子 B の間に働く力 分子 A と分子 B の間に働く力 のとき, A
 の気液平衡 はじめに 純物質 A( 液体 ) と純物質 B( 液体 ) が存在し, 分子 A の間に働く力 分子 B の間に働く力 分子 A と分子 B の間に働く力 のとき, A") との混合物 ( 二成分系 ) の気液平衡 はじめに 純物質 ( ) と純物質 ( ) が存在し, 分子 の間に働く力 分子 の間に働く力 分子 と分子 の間に働く力 のとき, と の混合物は任意の組成 ( モル分率 ) においてラウールの法則が成り立つ ラウールの法則 ある温度で純物質 が気液平衡状態にあるときの の蒸気圧 ( 飽和蒸気圧 ) を, 同温の を含む溶液が気液平衡状態にあるときの溶液中の
との混合物 ( 二成分系 ) の気液平衡 はじめに 純物質 ( ) と純物質 ( ) が存在し, 分子 の間に働く力 分子 の間に働く力 分子 と分子 の間に働く力 のとき, と の混合物は任意の組成 ( モル分率 ) においてラウールの法則が成り立つ ラウールの法則 ある温度で純物質 が気液平衡状態にあるときの の蒸気圧 ( 飽和蒸気圧 ) を, 同温の を含む溶液が気液平衡状態にあるときの溶液中の
我々のビッグデータ処理の新しい産業応用 広告やゲーム レコメンだけではない 個別化医療 ( ライフサイエンス ): 精神神経系疾患 ( うつ病 総合失調症 ) の網羅的ゲノム診断法の開発 全人類のゲノム解析と個別化医療実現を目標 ゲノム育種 ( グリーンサイエンス ): ブルーベリー オオムギ イネ
: 精神神経系疾患 ( うつ病 総合失調症 ) の網羅的ゲノム診断法の開発 全人類のゲノム解析と個別化医療実現を目標 ゲノム育種 ( グリーンサイエンス ): ブルーベリー オオムギ イネ") モンテカルロ法による分子進化の分岐図作成 のための最適化法 石井一夫 1 松田朋子 2 古崎利紀 1 後藤哲雄 2 1 東京農工大学 2 茨城大学 2013 9 9 2013 1 我々のビッグデータ処理の新しい産業応用 広告やゲーム レコメンだけではない 個別化医療 ( ライフサイエンス ): 精神神経系疾患 ( うつ病 総合失調症 ) の網羅的ゲノム診断法の開発 全人類のゲノム解析と個別化医療実現を目標
モンテカルロ法による分子進化の分岐図作成 のための最適化法 石井一夫 1 松田朋子 2 古崎利紀 1 後藤哲雄 2 1 東京農工大学 2 茨城大学 2013 9 9 2013 1 我々のビッグデータ処理の新しい産業応用 広告やゲーム レコメンだけではない 個別化医療 ( ライフサイエンス ): 精神神経系疾患 ( うつ病 総合失調症 ) の網羅的ゲノム診断法の開発 全人類のゲノム解析と個別化医療実現を目標
生理学 1章 生理学の基礎 1-1. 細胞の主要な構成成分はどれか 1 タンパク質 2 ビタミン 3 無機塩類 4 ATP 第5回 按マ指 (1279) 1-2. 細胞膜の構成成分はどれか 1 無機りん酸 2 リボ核酸 3 りん脂質 4 乳酸 第6回 鍼灸 (1734) E L 1-3. 細胞膜につ
 1-2. 細胞膜の構成成分はどれか 1 無機りん酸 2 リボ核酸 3 りん脂質 4 乳酸 第6回 鍼灸 (1734) E L 1-3. 細胞膜につ") の基礎 1-1. 細胞の主要な構成成分はどれか 1 タンパク質 2 ビタミン 3 無機塩類 4 ATP 第5回 (1279) 1-2. 細胞膜の構成成分はどれか 1 無機りん酸 2 リボ核酸 3 りん脂質 4 乳酸 第6回 (1734) 1-3. 細胞膜について正しい記述はどれか 1 糖脂質分子が規則正しく配列している 2 イオンに対して選択的な透過性をもつ 3 タンパク質分子の二重層膜からなる 4
の基礎 1-1. 細胞の主要な構成成分はどれか 1 タンパク質 2 ビタミン 3 無機塩類 4 ATP 第5回 (1279) 1-2. 細胞膜の構成成分はどれか 1 無機りん酸 2 リボ核酸 3 りん脂質 4 乳酸 第6回 (1734) 1-3. 細胞膜について正しい記述はどれか 1 糖脂質分子が規則正しく配列している 2 イオンに対して選択的な透過性をもつ 3 タンパク質分子の二重層膜からなる 4
本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因
 HbA1c 測定系について ~ 原理と特徴 ~ 一般社団法人日本臨床検査薬協会 技術運営委員会副委員長 安部正義 本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因 HPLC 法 HPLC 法原理 高速液体クロマトグラフィー 混合物の分析法の一つ 固体または液体の固定相 ( 吸着剤 ) 中で 液体または気体の移動相 ( 展開剤 ) に試料を加えて移動させ
HbA1c 測定系について ~ 原理と特徴 ~ 一般社団法人日本臨床検査薬協会 技術運営委員会副委員長 安部正義 本日の内容 HbA1c 測定方法別原理と特徴 HPLC 法 免疫法 酵素法 原理差による測定値の乖離要因 HPLC 法 HPLC 法原理 高速液体クロマトグラフィー 混合物の分析法の一つ 固体または液体の固定相 ( 吸着剤 ) 中で 液体または気体の移動相 ( 展開剤 ) に試料を加えて移動させ
細胞の構造
 大阪電気通信大学 5/8/18 本日の講義の内容 酵素 教科書 第 4 章 触媒反応とエネルギーの利用 酵素の性質 酵素反応の調節 酵素の種類 触媒の種類 無機物からなる無機触媒と有機物からなる有機触媒がある 触媒反応とエネルギーの利用 1 無機触媒の例 過酸化水素水に二酸化マンガンを入れると過酸化水素水が分解して水と酸素になる 2 有機触媒の例 細胞内に含まれるカタラーゼという酵素を過酸化水素水に加えると
大阪電気通信大学 5/8/18 本日の講義の内容 酵素 教科書 第 4 章 触媒反応とエネルギーの利用 酵素の性質 酵素反応の調節 酵素の種類 触媒の種類 無機物からなる無機触媒と有機物からなる有機触媒がある 触媒反応とエネルギーの利用 1 無機触媒の例 過酸化水素水に二酸化マンガンを入れると過酸化水素水が分解して水と酸素になる 2 有機触媒の例 細胞内に含まれるカタラーゼという酵素を過酸化水素水に加えると
アルゴリズム入門
 アルゴリズム入門 第 11 回 ~ パターン認識 (1)~ 情報理工学系研究科 創造情報学専攻 中山英樹 1 今日の内容 パターン認識問題の 1 つ : アラインメント アルゴリズム 再帰 動的計画法 2 パターン認識 音や画像の中に隠れたパターンを認識する 音素 音節 単語 文 基本図形 文字 指紋 物体 人物 顔 パターン は唯一のデータではなく 似通ったデータの集まりを表している 多様性 ノイズ
アルゴリズム入門 第 11 回 ~ パターン認識 (1)~ 情報理工学系研究科 創造情報学専攻 中山英樹 1 今日の内容 パターン認識問題の 1 つ : アラインメント アルゴリズム 再帰 動的計画法 2 パターン認識 音や画像の中に隠れたパターンを認識する 音素 音節 単語 文 基本図形 文字 指紋 物体 人物 顔 パターン は唯一のデータではなく 似通ったデータの集まりを表している 多様性 ノイズ
記者発表資料
 2012 年 6 月 4 日 報道機関各位 東北大学流体科学研究所原子分子材料科学高等研究機構 高密度 均一量子ナノ円盤アレイ構造による高効率 量子ドット太陽電池の実現 ( シリコン量子ドット太陽電池において世界最高変換効率 12.6% を達成 ) < 概要 > 東北大学 流体科学研究所および原子分子材料科学高等研究機構 寒川教授グループはこの度 新しい鉄微粒子含有蛋白質 ( リステリアフェリティン
2012 年 6 月 4 日 報道機関各位 東北大学流体科学研究所原子分子材料科学高等研究機構 高密度 均一量子ナノ円盤アレイ構造による高効率 量子ドット太陽電池の実現 ( シリコン量子ドット太陽電池において世界最高変換効率 12.6% を達成 ) < 概要 > 東北大学 流体科学研究所および原子分子材料科学高等研究機構 寒川教授グループはこの度 新しい鉄微粒子含有蛋白質 ( リステリアフェリティン
ナノの技術をバイオに応用
 本日まで お試し期間 なので 出席は取りません 現代生物学概論 2 遺伝子 ( プログラム ) と蛋白質 ( ナノマシン ) 先進理工学科 化学生物学研究室 准教授 生体機能システムコース 瀧真清 1 本日の概要 : 蛋白質生合成の全スキーム D から蛋白質への情報の流れ アミノ酸から蛋白質への物質の流れ 転写 D 本日は詳細は省略 アミノアシル tr 合成酵素 (RS) 翻訳 mr コドンーアンチコドンの対合
本日まで お試し期間 なので 出席は取りません 現代生物学概論 2 遺伝子 ( プログラム ) と蛋白質 ( ナノマシン ) 先進理工学科 化学生物学研究室 准教授 生体機能システムコース 瀧真清 1 本日の概要 : 蛋白質生合成の全スキーム D から蛋白質への情報の流れ アミノ酸から蛋白質への物質の流れ 転写 D 本日は詳細は省略 アミノアシル tr 合成酵素 (RS) 翻訳 mr コドンーアンチコドンの対合
PowerPoint プレゼンテーション
 細胞の情報伝達 (1) 何を学習するか細胞が環境からシグナル ( 刺激 ) を受けて 細胞の状態が変化するときに 細胞内でどのような現象が起きているか を知る分子の大変複雑な連続反応であるので 反応の最初の段階を中心に見ていく ( 共通の現象が多いから ; 疾患の治療の標的となる分子が多い ) これを知るために (2) リガンドの拡散様式 ( 図 16-3) リガンドを発現する細胞とこれを受け取る細胞との
細胞の情報伝達 (1) 何を学習するか細胞が環境からシグナル ( 刺激 ) を受けて 細胞の状態が変化するときに 細胞内でどのような現象が起きているか を知る分子の大変複雑な連続反応であるので 反応の最初の段階を中心に見ていく ( 共通の現象が多いから ; 疾患の治療の標的となる分子が多い ) これを知るために (2) リガンドの拡散様式 ( 図 16-3) リガンドを発現する細胞とこれを受け取る細胞との
Microsoft PowerPoint _生物配列解析基礎_3回目.pptx
 1 古細菌 真正細菌 3 4 多くの生物にはDNA修復を行う機 構が備わっており これらをDNA 突然変異 修復系と呼ぶ アルビノのカラス 1つのDNAに生じた突然変異によって鎌状赤血球貧血症になる 5 進化の総合説 現在 進化を説明する理論 として最も支持されている のは進化の総合説と呼ばれ るもので 自然選択説や突 然変異説 隔離説 メンデ ルの遺伝子の理論 集団遺 伝学の理論や中立進化説な どを統合したものである
1 古細菌 真正細菌 3 4 多くの生物にはDNA修復を行う機 構が備わっており これらをDNA 突然変異 修復系と呼ぶ アルビノのカラス 1つのDNAに生じた突然変異によって鎌状赤血球貧血症になる 5 進化の総合説 現在 進化を説明する理論 として最も支持されている のは進化の総合説と呼ばれ るもので 自然選択説や突 然変異説 隔離説 メンデ ルの遺伝子の理論 集団遺 伝学の理論や中立進化説な どを統合したものである
GWB
 NGS データ解析入門 Web セミナー : De Novo シークエンス解析編 1 NGS 新規ゲノム配列解析の手順 シークエンス 遺伝子領域の検出 アセンブル データベース検索 2 解析ワークフローと使用ソフトウェア シークエンスデータのインポート クオリティチェック 前処理 コンティグ配列の作成 CLC Genomics Workbench 遺伝子領域の検出 Blast2GO PRO データベース検索
NGS データ解析入門 Web セミナー : De Novo シークエンス解析編 1 NGS 新規ゲノム配列解析の手順 シークエンス 遺伝子領域の検出 アセンブル データベース検索 2 解析ワークフローと使用ソフトウェア シークエンスデータのインポート クオリティチェック 前処理 コンティグ配列の作成 CLC Genomics Workbench 遺伝子領域の検出 Blast2GO PRO データベース検索
Gifu University Faculty of Engineering
 Gifu University Faculty of Engineering Gifu University Faculty of Engineering the structure of the faculty of engineering DATA Gifu University Faculty of Engineering the aim of the university education
Gifu University Faculty of Engineering Gifu University Faculty of Engineering the structure of the faculty of engineering DATA Gifu University Faculty of Engineering the aim of the university education
34 タンパク質の立体構造予測 タンパク質の配列から 立体構造を予測します あらゆるレベルに応じた対応をします モデリング可能なもの高精度モデリングを行います モデリングが難しいもの技術の粋をこらして鋳型を探します 人工鋳型も作成します 構造がないもの天然変性領域を予測します 支援に供する設備名など
 33 MD-SAXS 法 マルチドメインタンパク質や天然変性タンパク質など フレキシブルで結晶化しにくく X 線結晶構造解析が難しいタンパク質は数多く存在する また 結晶構造と溶液構造が異なると想定される場合もある そのような場合 低解像度ながら 溶液構造情報を X 線小角散乱 (SAXS) 実験で得ることができる そこで バイオインフォマティクス技術を使ってモデリングした立体構造に対し 分子動力学
33 MD-SAXS 法 マルチドメインタンパク質や天然変性タンパク質など フレキシブルで結晶化しにくく X 線結晶構造解析が難しいタンパク質は数多く存在する また 結晶構造と溶液構造が異なると想定される場合もある そのような場合 低解像度ながら 溶液構造情報を X 線小角散乱 (SAXS) 実験で得ることができる そこで バイオインフォマティクス技術を使ってモデリングした立体構造に対し 分子動力学
共起関係解析によるタンパク質の機能モジュール探索法の開発
 2014.03.02 統合化推進プログラム統合データ解析トライアル研究成果報告会 共起関係解析によるタンパク質の機能モジュール探索法の開発 九州工業大学 情報工 生命情報 藤井聡 2014 藤井聡 ( 九州工業大学 )licensed under CC 表示 2.1 日本 1 背景 疾病関連遺伝子 ドメイン モチーフ ドラッグターゲット T/G Gefitinib LUNG CANCER OMIM
2014.03.02 統合化推進プログラム統合データ解析トライアル研究成果報告会 共起関係解析によるタンパク質の機能モジュール探索法の開発 九州工業大学 情報工 生命情報 藤井聡 2014 藤井聡 ( 九州工業大学 )licensed under CC 表示 2.1 日本 1 背景 疾病関連遺伝子 ドメイン モチーフ ドラッグターゲット T/G Gefitinib LUNG CANCER OMIM
PowerPoint プレゼンテーション
 酵素 : タンパク質の触媒 タンパク質 Protein 酵素 Enzyme 触媒 Catalyst 触媒 Cataylst: 特定の化学反応の反応速度を速める物質 自身は反応の前後で変化しない 酵素 Enzyme: タンパク質の触媒 触媒作用を持つタンパク質 第 3 回 : タンパク質はアミノ酸からなるポリペプチドである 第 4 回 : タンパク質は様々な立体構造を持つ 第 5 回 : タンパク質の立体構造と酵素活性の関係
酵素 : タンパク質の触媒 タンパク質 Protein 酵素 Enzyme 触媒 Catalyst 触媒 Cataylst: 特定の化学反応の反応速度を速める物質 自身は反応の前後で変化しない 酵素 Enzyme: タンパク質の触媒 触媒作用を持つタンパク質 第 3 回 : タンパク質はアミノ酸からなるポリペプチドである 第 4 回 : タンパク質は様々な立体構造を持つ 第 5 回 : タンパク質の立体構造と酵素活性の関係
Microsoft PowerPoint - 04_01_text_UML_03-Sequence-Com.ppt
 システム設計 (1) シーケンス図 コミュニケーション図等 1 今日の演習のねらい 2 今日の演習のねらい 情報システムを構成するオブジェクトの考え方を理解す る 業務プロセスでのオブジェクトの相互作用を考える シーケンス図 コミュニケーション図を作成する 前回までの講義システム開発の上流工程として 要求仕様を確定パソコンを注文するまでのユースケースユースケースから画面の検討イベントフロー アクティビティ図
システム設計 (1) シーケンス図 コミュニケーション図等 1 今日の演習のねらい 2 今日の演習のねらい 情報システムを構成するオブジェクトの考え方を理解す る 業務プロセスでのオブジェクトの相互作用を考える シーケンス図 コミュニケーション図を作成する 前回までの講義システム開発の上流工程として 要求仕様を確定パソコンを注文するまでのユースケースユースケースから画面の検討イベントフロー アクティビティ図
脳 -分子・遺伝子・生理- (立ち読み)
") i Brain and eural Functions by SII ISIUA BU SASAGAWA EUGEE FUTAI 電子書籍の不正コピーは法律により罰せられます 本作品の著作権その他の法的権利は 本作品の著作者ならびに裳華房その他第三者に帰属します 本作品の全部または一部について 権利者に無断で 複製 公衆送信 出版 貸与 翻訳 翻案および改編するなど 本作品の権利を侵害する方法で利用することを禁止します
i Brain and eural Functions by SII ISIUA BU SASAGAWA EUGEE FUTAI 電子書籍の不正コピーは法律により罰せられます 本作品の著作権その他の法的権利は 本作品の著作者ならびに裳華房その他第三者に帰属します 本作品の全部または一部について 権利者に無断で 複製 公衆送信 出版 貸与 翻訳 翻案および改編するなど 本作品の権利を侵害する方法で利用することを禁止します
ヒトゲノム情報を用いた創薬標的としての新規ペプチドリガンドライブラリー PharmaGPEP TM Ver2S のご紹介 株式会社ファルマデザイン
 ヒトゲノム情報を用いた創薬標的としての新規ペプチドリガンドライブラリー PharmaGPEP TM Ver2S のご紹介 株式会社ファルマデザイン 薬剤の標的分子別構成 核内受容体 2% DNA 2% ホルモン 成長因子 11% 酵素 28% イオンチャンネル 5% その他 7% 受容体 45% Drews J,Science 287,1960-1964(2000) G 蛋白質共役受容体 (GPCR)
ヒトゲノム情報を用いた創薬標的としての新規ペプチドリガンドライブラリー PharmaGPEP TM Ver2S のご紹介 株式会社ファルマデザイン 薬剤の標的分子別構成 核内受容体 2% DNA 2% ホルモン 成長因子 11% 酵素 28% イオンチャンネル 5% その他 7% 受容体 45% Drews J,Science 287,1960-1964(2000) G 蛋白質共役受容体 (GPCR)
PowerPoint Presentation
 パターン認識入門 パターン認識 音や画像に中に隠れたパターンを認識する 音素 音節 単語 文 基本図形 文字 指紋 物体 人物 顔 パターン は唯一のデータではなく 似通ったデータの集まりを表している 多様性 ノイズ 等しい から 似ている へ ~ だ から ~ らしい へ 等しい から 似ている へ 完全に等しいかどうかではなく 似ているか どうかを判定する パターンを代表する模範的データとどのくらい似ているか
パターン認識入門 パターン認識 音や画像に中に隠れたパターンを認識する 音素 音節 単語 文 基本図形 文字 指紋 物体 人物 顔 パターン は唯一のデータではなく 似通ったデータの集まりを表している 多様性 ノイズ 等しい から 似ている へ ~ だ から ~ らしい へ 等しい から 似ている へ 完全に等しいかどうかではなく 似ているか どうかを判定する パターンを代表する模範的データとどのくらい似ているか
 NCBI BLAST チュートリアル このチュートリアルでは NCBI サイトでの BLAST による相同性検索の方法について 一般的な使い方を紹介しています はじめに. BLAST とは まずはじめに 簡単に BLAST について紹介することにしましょう BLAST は Basic Local Alignment Search Tool の略で ペアワイズの局所的なアライメント / 相同性検索 (
NCBI BLAST チュートリアル このチュートリアルでは NCBI サイトでの BLAST による相同性検索の方法について 一般的な使い方を紹介しています はじめに. BLAST とは まずはじめに 簡単に BLAST について紹介することにしましょう BLAST は Basic Local Alignment Search Tool の略で ペアワイズの局所的なアライメント / 相同性検索 (
<4D F736F F F696E74202D2091E682508FCD E836D8E5F82C6835E E8EBF205B8CDD8AB B83685D>
 生物有機化学 教科書 : マクマリー 生物有機化学生化学編 rganic and biological chemistrybiochemistry 生物 ( 生命 ) をより深く理解する 分子レベルで = 有機化学的に 地球の凄さ水 + 空気の世界 ( 2, 2 ) 水 : 酵素反応の溶媒 ( 反応の場を提供 ) ものを溶かす ( 物質によって溶解性は違う ) Why? 水 : 酵素反応の溶媒 (
生物有機化学 教科書 : マクマリー 生物有機化学生化学編 rganic and biological chemistrybiochemistry 生物 ( 生命 ) をより深く理解する 分子レベルで = 有機化学的に 地球の凄さ水 + 空気の世界 ( 2, 2 ) 水 : 酵素反応の溶媒 ( 反応の場を提供 ) ものを溶かす ( 物質によって溶解性は違う ) Why? 水 : 酵素反応の溶媒 (
7-1(DNA配列から遺伝子を探す).ppt
.ppt") DNA 配列の中から遺伝子を探す Blast 解析.6 Query DNA 塩基配列アミノ酸配列 DNA 塩基配列をアミノ酸配列に変換アミノ酸配列 DNA 塩基配列をアミノ酸配列に変換 データベース DNA 塩基配列アミノ酸配列アミノ酸配列 DNA 塩基配列をアミノ酸配列に変換 DNA 塩基配列をアミノ酸配列に変換 1. 2. 3. TATGGCTTA---- T G L TATGGCTTA----
DNA 配列の中から遺伝子を探す Blast 解析.6 Query DNA 塩基配列アミノ酸配列 DNA 塩基配列をアミノ酸配列に変換アミノ酸配列 DNA 塩基配列をアミノ酸配列に変換 データベース DNA 塩基配列アミノ酸配列アミノ酸配列 DNA 塩基配列をアミノ酸配列に変換 DNA 塩基配列をアミノ酸配列に変換 1. 2. 3. TATGGCTTA---- T G L TATGGCTTA----
研究成果報告書
 PHF (paired helical filament) Tau2r3 Fab2r3 MBD 1 MBD (1) The microtubule-binding domain (MBD) of tau protein is the region responsible for the PHF formation. We succeed in the preparation of an antibody
PHF (paired helical filament) Tau2r3 Fab2r3 MBD 1 MBD (1) The microtubule-binding domain (MBD) of tau protein is the region responsible for the PHF formation. We succeed in the preparation of an antibody
創薬に繋がる V-ATPase の構造 機能の解明 Towards structure-based design of novel inhibitors for V-ATPase 京都大学医学研究科 / 理化学研究所 SSBC 村田武士 < 要旨 > V-ATPase は 真核生物の空胞系膜に存在す
 創薬に繋がる V-ATPase の構造 機能の解明 Towards structure-based design of novel inhibitors for V-ATPase 京都大学医学研究科 / 理化学研究所 SSBC 村田武士 < 要旨 > V-ATPase は 真核生物の空胞系膜に存在するプロトンポンプである 複雑なサブユニット構造からなる超分子複合体であり 親水性の触媒頭部部分 (V1
創薬に繋がる V-ATPase の構造 機能の解明 Towards structure-based design of novel inhibitors for V-ATPase 京都大学医学研究科 / 理化学研究所 SSBC 村田武士 < 要旨 > V-ATPase は 真核生物の空胞系膜に存在するプロトンポンプである 複雑なサブユニット構造からなる超分子複合体であり 親水性の触媒頭部部分 (V1
Microsoft Word - PRESS_
 ニュースリリース 平成 20 年 8 月 1 日千葉大学大学院園芸学研究科 新たな基盤転写 (RNA 合成 ) 系の発見 原始生物シゾンで解明されたリボゾーム RNA 合成系進化のミッシングリンク < 研究成果の概要 > 本学園芸学研究科の田中寛教授 今村壮輔 JSPS 特別研究員 華岡光正東京大学研究員は 植物に残されていた始原的なリボゾーム RNA 合成系を発見し これまで不明だったリボゾーム
ニュースリリース 平成 20 年 8 月 1 日千葉大学大学院園芸学研究科 新たな基盤転写 (RNA 合成 ) 系の発見 原始生物シゾンで解明されたリボゾーム RNA 合成系進化のミッシングリンク < 研究成果の概要 > 本学園芸学研究科の田中寛教授 今村壮輔 JSPS 特別研究員 華岡光正東京大学研究員は 植物に残されていた始原的なリボゾーム RNA 合成系を発見し これまで不明だったリボゾーム
PowerPoint プレゼンテーション
 機能アノテーションパイプライン ( 仮 ) 理化学研究所発生 再生科学総合研究センター (CDB) 機能ゲノミクスサブユニット粕川雄也 1 発表の概要 機能アノテーションってなに? 機能アノテーションはどうやってつけるの? パイプライン化 & ハイスループット化するには? 2 発表の概要 機能アノテーションってなに? 機能アノテーションはどうやってつけるの? パイプライン化 & ハイスループット化するには?
機能アノテーションパイプライン ( 仮 ) 理化学研究所発生 再生科学総合研究センター (CDB) 機能ゲノミクスサブユニット粕川雄也 1 発表の概要 機能アノテーションってなに? 機能アノテーションはどうやってつけるの? パイプライン化 & ハイスループット化するには? 2 発表の概要 機能アノテーションってなに? 機能アノテーションはどうやってつけるの? パイプライン化 & ハイスループット化するには?
線形代数とは
 線形代数とは 第一回ベクトル 教科書 エクササイズ線形代数 立花俊一 成田清正著 共立出版 必要最低限のことに限る 得意な人には物足りないかもしれません 線形代数とは何をするもの? 線形関係 y 直線 yもも 次式で登場する (( 次の形 ) 線形 ただし 次元の話世の中は 3 次元 [4[ 次元 ] 次元 3 次元 4 次元 はどうやって直線を表すの? ベクトルや行列の概念 y A ベクトルを使うと
線形代数とは 第一回ベクトル 教科書 エクササイズ線形代数 立花俊一 成田清正著 共立出版 必要最低限のことに限る 得意な人には物足りないかもしれません 線形代数とは何をするもの? 線形関係 y 直線 yもも 次式で登場する (( 次の形 ) 線形 ただし 次元の話世の中は 3 次元 [4[ 次元 ] 次元 3 次元 4 次元 はどうやって直線を表すの? ベクトルや行列の概念 y A ベクトルを使うと
2. 看護に必要な栄養と代謝について説明できる 栄養素としての糖質 脂質 蛋白質 核酸 ビタミンなどの性質と役割 およびこれらの栄養素に関連する生命活動について具体例を挙げて説明できる 生体内では常に物質が交代していることを説明できる 代謝とは エネルギーを生み出し 生体成分を作り出す反応であること
 生化学 責任者 コーディネーター 看護専門基礎講座塚本恭正准教授 担当講座 学科 ( 分野 ) 看護専門基礎講座 対象学年 1 期間後期 区分 時間数 講義 22.5 時間 単位数 2 単位 学習方針 ( 講義概要等 ) 生化学反応の場となる細胞と細胞小器官の構造と機能を理解する エネルギー ATP を産生し 生体成分を作り出す代謝反応が生命活動で果たす役割を理解し 代謝反応での酵素の働きを学ぶ からだを構成する蛋白質
生化学 責任者 コーディネーター 看護専門基礎講座塚本恭正准教授 担当講座 学科 ( 分野 ) 看護専門基礎講座 対象学年 1 期間後期 区分 時間数 講義 22.5 時間 単位数 2 単位 学習方針 ( 講義概要等 ) 生化学反応の場となる細胞と細胞小器官の構造と機能を理解する エネルギー ATP を産生し 生体成分を作り出す代謝反応が生命活動で果たす役割を理解し 代謝反応での酵素の働きを学ぶ からだを構成する蛋白質
Untitled
 上原記念生命科学財団研究報告集, 25 (2011) 86. 線虫 C. elegans およびマウスをモデル動物とした体細胞レベルで生じる性差の解析 井上英樹 Key words: 性差, ストレス応答,DMRT 立命館大学生命科学部生命医科学科 緒言性差は雌雄の性に分かれた動物にみられ, 生殖能力の違いだけでなく形態, 行動などそれぞれの性の間でみられる様々な差異と定義される. 性差は, 形態や行動だけでなく疾患の発症リスクの男女差といった生理的なレベルの差異も含まれる.
上原記念生命科学財団研究報告集, 25 (2011) 86. 線虫 C. elegans およびマウスをモデル動物とした体細胞レベルで生じる性差の解析 井上英樹 Key words: 性差, ストレス応答,DMRT 立命館大学生命科学部生命医科学科 緒言性差は雌雄の性に分かれた動物にみられ, 生殖能力の違いだけでなく形態, 行動などそれぞれの性の間でみられる様々な差異と定義される. 性差は, 形態や行動だけでなく疾患の発症リスクの男女差といった生理的なレベルの差異も含まれる.
先端生物工学演習Ⅱ 「タンパク質の電気泳動」
 先端生物工学演習 Ⅱ タンパク質の電気泳動 2008 年 10 月 14 日 ( 旧 ) 進化生命システム学塚田幸治 1 話の中身 電気泳動という手法 ( 一般 ) アミノ酸の電荷とタンパク質の電荷 具体的な実験手法について ( 原理 ) Native( ( 構造や機能が破壊されていないタンパク質 ) SDS( ( 変性条件下でのタンパク質 ) 応用例 等電点電気泳動法と二次元電気泳動 非特異的染色法と特異的染色法
先端生物工学演習 Ⅱ タンパク質の電気泳動 2008 年 10 月 14 日 ( 旧 ) 進化生命システム学塚田幸治 1 話の中身 電気泳動という手法 ( 一般 ) アミノ酸の電荷とタンパク質の電荷 具体的な実験手法について ( 原理 ) Native( ( 構造や機能が破壊されていないタンパク質 ) SDS( ( 変性条件下でのタンパク質 ) 応用例 等電点電気泳動法と二次元電気泳動 非特異的染色法と特異的染色法
KEGG.ppt
 1 2 3 4 KEGG: Kyoto Encyclopedia of Genes and Genomes http://www.genome.jp/kegg/kegg2.html http://www.genome.jp/kegg/kegg_ja.html 5 KEGG PATHWAY 生体内(外)の分子間ネットワーク図 代謝系 12カテゴリ 中間代謝 二次代謝 薬の 代謝 全体像 制御系 20カテゴリ
1 2 3 4 KEGG: Kyoto Encyclopedia of Genes and Genomes http://www.genome.jp/kegg/kegg2.html http://www.genome.jp/kegg/kegg_ja.html 5 KEGG PATHWAY 生体内(外)の分子間ネットワーク図 代謝系 12カテゴリ 中間代謝 二次代謝 薬の 代謝 全体像 制御系 20カテゴリ
オートマトン 形式言語及び演習 1. 有限オートマトンとは 酒井正彦 形式言語 言語とは : 文字列の集合例 : 偶数個の 1 の後に 0 を持つ列からなる集合 {0, 110, 11110,
 オートマトン 形式言語及び演習 1 有限オートマトンとは 酒井正彦 wwwtrscssinagoya-uacjp/~sakai/lecture/automata/ 形式言語 言語とは : 文字列の集合例 : 偶数個の 1 の後に 0 を持つ列からなる集合 {0, 110, 11110, } 形式言語 : 数学モデルに基づいて定義された言語 認識機械 : 文字列が該当言語に属するか? 文字列 機械 受理
オートマトン 形式言語及び演習 1 有限オートマトンとは 酒井正彦 wwwtrscssinagoya-uacjp/~sakai/lecture/automata/ 形式言語 言語とは : 文字列の集合例 : 偶数個の 1 の後に 0 を持つ列からなる集合 {0, 110, 11110, } 形式言語 : 数学モデルに基づいて定義された言語 認識機械 : 文字列が該当言語に属するか? 文字列 機械 受理
研修コーナー
 l l l l l l l l l l l α α β l µ l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l
l l l l l l l l l l l α α β l µ l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l l
<4D6963726F736F667420506F776572506F696E74202D203134303333305F945F8C7C89BB8A7789EF83568393837C8357834583808169938C91E5814589699363816A8CF68A4A97702E707074205B8CDD8AB78382815B83685D>
 構 造 生 物 学 とバイオインフォマティクスの 境 界 領 域 における 研 究 支 援 と 高 度 化 創 薬 等 PF 情 報 拠 点 東 大 院 農 生 科 永 田 宏 次 情 報 拠 点 分 担 課 題 : 構 造 生 物 学 的 立 場 からのデータベース 運 用 支 援 と 高 度 化 情 報 領 域 東 京 大 学 ( 永 田 宏 次 ) 課 題 概 要 全 国 の 生 命 科 学
構 造 生 物 学 とバイオインフォマティクスの 境 界 領 域 における 研 究 支 援 と 高 度 化 創 薬 等 PF 情 報 拠 点 東 大 院 農 生 科 永 田 宏 次 情 報 拠 点 分 担 課 題 : 構 造 生 物 学 的 立 場 からのデータベース 運 用 支 援 と 高 度 化 情 報 領 域 東 京 大 学 ( 永 田 宏 次 ) 課 題 概 要 全 国 の 生 命 科 学
60 秒でわかるプレスリリース 2008 年 10 月 20 日 独立行政法人理化学研究所 アルツハイマー病の原因となる アミロイドベータ の産生調節機構を解明 - 新しいアルツハイマー病治療薬の開発に有望戦略 - 高年齢化社会を迎え 認知症に対する対策が社会的な課題となっています 国内では 認知症
 60 秒でわかるプレスリリース 2008 年 10 月 20 日 独立行政法人理化学研究所 アルツハイマー病の原因となる アミロイドベータ の産生調節機構を解明 - 新しいアルツハイマー病治療薬の開発に有望戦略 - 高年齢化社会を迎え 認知症に対する対策が社会的な課題となっています 国内では 認知症老人が約 100 万人を超え その約半分はアルツハイマー病患者とされています その数はさらに増え続けると予想され
60 秒でわかるプレスリリース 2008 年 10 月 20 日 独立行政法人理化学研究所 アルツハイマー病の原因となる アミロイドベータ の産生調節機構を解明 - 新しいアルツハイマー病治療薬の開発に有望戦略 - 高年齢化社会を迎え 認知症に対する対策が社会的な課題となっています 国内では 認知症老人が約 100 万人を超え その約半分はアルツハイマー病患者とされています その数はさらに増え続けると予想され